Visione a colori per mezzo di reti neurali
Frontiere della Vita (1999)
Visione a colori per mezzo di reti neurali
In questo saggio mostreremo come la comprensione delle funzioni interne cerebrali possa essere migliorata da una rappresentazione basata su reti neurali artificiali, combinando insieme strategie di tipo top-down e bottom-up. Determinate funzioni elaborative del sistema nervoso vengono usate per addestrare le reti, e le rappresentazioni interne di queste vengono in seguito esaminate e confrontate con i dati sperimentali tratti da singoli neuroni o da funzioni cerebrali reali. Metteremo in luce come, attraverso processi di apprendimento supervisionato e non supervisionato, venga acquisita una rappresentazione interna del colore nel sistema visivo, analizzando in particolare le relazioni tra le sequenze di attivazione e le funzionalità di apprendimento. Illustreremo come le reti neurali non servano solo a riprodurre artificialmente una data funzionalità: attraverso l'esame delle rappresentazioni interne, specifiche per una certa particolare funzionalità, esse forniscono anche informazioni su ciò che avviene, e con quali modalità, nei sistemi nervosi reali.
Introduzione
Alcuni recenti progressi nello studio fisiologico della visione a colori hanno rivelato l'esistenza, nel percorso visivo, di diversi tipi di rappresentazione del colore. Sebbene molti modelli della visione a colori siano in grado di spiegare il modo in cui l'informazione cromatica viene trasformata lungo il percorso visivo, solo pochi tentano di spiegare i processi logici su cui si basa l'elaborazione di tale informazione. In questo saggio, dopo aver passato in rassegna le osservazioni sperimentali sulla fisiologia della visione a colori, affronteremo i problemi di tipo elaborativo legati alla rappresentazione cromatica lungo il percorso visivo, con particolare riferimento ai vantaggi insiti in tale rappresentazione. Successivamente descriveremo un modello della visione cromatica basato su reti neurali con capacità di apprendimento supervisionato e non supervisionato. Tale modello si dimostra utile, in particolare, in problemi di opponenza o di costanza cromatica e nella individuazione dei campi recettivi cromatici.
Rappresentazione del colore lungo il percorso visivo
Dati fisiologici
La visione a colori ha origine nei fotorecettori presenti nella retina, cioè nei coni, che sono di tre tipi differenti, ciascuno dei quali risponde selettivamente a regioni diverse dello spettro della luce visibile. La sensibilità spettrale dei coni è stata misurata mediante osservazioni psicofisiche, tecniche microspettrofotometriche ed esperimenti di elettrofisiologia, ottenendo un buon accordo nella maggior parte dello spettro. Queste misure hanno dimostrato che la sensibilità spettrale dei coni è ampia e sovrapposta, permettendo di considerare triplette di risposte dei coni come il primo stadio della rappresentazione del colore lungo il percorso visivo.
Molti studi hanno messo in evidenza che esiste uno stadio successivo a quello dei recettori, nel quale i segnali tricromatici provenienti dai coni vengono trasformati in segnali acromatici e a contrasto di colore. Attraverso studi psicofisici, si sono misurate le sensibilità spettrali dei meccanismi coinvolti in questo secondo stadio. Sebbene si siano trovate difficoltà nel determinare la natura precisa di questo secondo stadio di trasformazione a partire dalle sole osservazioni psicofisiche, questi esperimenti hanno dimostrato l' esistenza di meccanismi di opponenza cromatica nel percorso visivo. I primi esperimenti hanno registrato segnali derivanti dall' opponenza cromatica da parte delle cellule orizzontali della retina dei pesci, dalle cellule gangliari e dai neuroni del nucleo genicolato laterale (LGN, Lateral Geniculate Nucleus) del macaco (Lennie e D'Zmura, 1988).
Alcune prove recenti dell'esistenza del meccanismo di opponenza cromatica (Derrington et al., 1984) indicano che nell'LGN parvocellulare esistono gruppi ben distinti di neuroni per le coppie di colori rosso-verde e giallo-blu, e che più del 90% dei neuroni appartiene al tipo rossoverde. Si è anche dimostrato che la maggior parte dei neuroni di selettività cromatica nello strato 4Cβ dell'area VI, che sono i destinatari principali del segnale di input parvocellulare, ha la stessa ampia selettività cromatica dei neuroni presenti nell'LGN (Lennie et al., 1990). Alcuni neuroni dello strato 4Cβ inviano segnali a regioni localizzate dello strato 3 dell'area VI colorata con citocromoossidasi. Queste regioni simili a chiazze vengono chiamate blob, e questa caratteristica costituisce un aspetto peculiare degli strati 2 e 3 dell'area VI. M.S. Livingstone e D.H. Hubel (1984) hanno trovato che più del 50% dei neuroni nei blob hanno campi recettivi a doppio contrasto cromatico, il centro e la periferia dei quali ricevono i segnali di input da due tipi diversi di coni, con segno opposto. Sebbene vi siano delle discrepanze nell'individuazione dei campi recettivi a doppia opponenza cromatica, a causa dei diversi criteri usati per la loro classificazione, questo tipo di campi potrebbe avere un ruolo nei processi di elaborazione del contrasto di colore o nella rilevazione dei bordi di colore.
Le proprietà cromatiche dei neuroni della corteccia extrastriata mostrano, d'altra parte, caratteristiche alquanto diverse. S.M. Zeki (1980) ha trovato che i neuroni dell'area V 4 sono selettivamente sensibili a regioni ristrette dello spettro. Alcuni studi (Schein et al., 1982) indicano che i neuroni di quest'area non sono più selettivi di quelli appartenenti a livelli inferiori del percorso visivo. Ciò nonostante, i neuroni in V 4 si distinguono nettamente dagli altri poiché non mostrano sensibilità palese a segnali di opponenza cromatica, ma hanno campi recettivi di caratteristiche particolari (Schein e Desimone, 1990). Tali campi presentano, al centro, un andamento della sensibilità spettrale con uno o due picchi stretti, mentre, nelle parti periferiche, pur mostrando una sensibilità spettrale simile, hanno un comportamento fortemente inibitorio. Ciò potrebbe giocare un ruolo essenziale nell'individuazione della costanza cromatica, anche se non è molto chiaro come possa essere tenuto in conto il colore illuminante. Dagli esperimenti volti a studiare la sensibilità spettrale dei neuroni della corteccia inferotemporale (IT), che ricevono intensi segnali dall'area V 4, si è trovato che il 70% di essi sono selettivi al colore, e che le tonalità di colore preferenziali variano da cellula a cellula (Komatsu et al., 1992). Questi risultati sulla selettività al colore dei neuroni delle aree V4 e IT suggeriscono l'ipotesi che la rappresentazione del colore in queste aree sia molto diversa da quella nei livelli inferiori del percorso visivo, come le aree VI o LGN. Sembra che nella corteccia visiva superiore il colore sia rappresentato in un numero di dimensioni maggiore di tre.
Problemi di rappresentazione ed elaborazione dell'informazione cromatica
Le osservazioni di carattere fisiologico descritte finora dimostrano che sono presenti tre tipi di rappresentazione del colore nel percorso visivo: rappresentazione in tricromia a livello sensoriale, rappresentazione a opponenza cromatica a livello intermedio, e rappresentazione multidimensionaIe a livelli corti cali superiori. Tuttavia, dal punto di vista computazionale, sorgono i seguenti interrogativi: perché l'informazione cromatica è rappresentata in questo modo? In quale maniera tali forme di rappresentazione sono correlate agli obiettivi computazionali?
Dagli studi fisiologici e anatomici è noto che le immagini retiniche campionate dai tre tipi di coni sono trasmesse attraverso il nervo ottico. Tuttavia, poiché il numero di cellule gangliari (circa 1,5 milioni) è minore di quello dei fotorecettori (circa 6 milioni) e l'intervallo dinamico proprio del nervo ottico è minore di quello dei fotorecettori, le immagini retiniche trasmesse risultano degradate. Tale degradazione è tuttavia ammissibile, e anche conveniente, per la presenza di correlazione interna al segnale trasmesso. Sono infatti presenti correlazioni spaziali derivanti dagli stimoli provenienti dall' ambiente naturale e correlazioni cromati che derivanti dalle risposte dei tre tipi di coni, la cui sensibilità spettrale è in buona parte sovrapposta. Si può pertanto supporre che il sistema visivo sia realizzato in modo ottimale per elaborare segnali molto correlati, o ridondanti, e sia specializzato nella costruzione di rappresentazioni efficienti del colore. Partendo da tale punto di vista, nei prossimi paragrafi introdurremo modelli computazionali della visione a colori, per mezzo dei quali proporremo giustificazioni delle varie forme di rappresentazione cromatica. Tali modelli saranno basati sulle reti neurali, con particolare riferimento alle loro capacità di apprendimento. È infatti prevedibile che tali reti rappresentino dei mezzi efficienti per interpretare dal punto di vista computazionale i dati psicologici e psicofisici, in quanto sia le opportune finalità computazionali, sia i prefissati criteri di ottimizzazione possono essere inseriti nella procedura di apprendimento. In questo modo si generano rappresentazioni interne in elementi funzionali operanti in parallelo, proprio come avviene nel cervello.
Modelli di visione a colori basati su reti neurali con apprendimento supervisionato
Rappresentazioni interne e apprendimento supervisionato
Il più diffuso metodo di apprendimento supervisionato per reti neurali multi strato è l'algoritrno di retropropagazione (backpropagation), che agisce secondo il criterio di minimizzazione dell'errore tra i segnali di output del modello e i segnali esterni di addestramento. Le reti basate su tale algoritmo possono fornire uno strumento potente per generare relazioni di corrispondenza non lineare, e sono state applicate con successo in problemi ingegneristici, quali il riconoscimento del parlato o dei caratteri manoscritti. In aggiunta a tali applicazioni, è tuttavia possibile mostrare che reti neurali basate sulla retropropagazione consentono di creare modelli funzionali, a livello di sistema, di parti specifiche del cervello. Una caratteristica interessante di tali reti risiede nello schema di attivazione delle unità dello strato nascosto, le quali, da alcuni punti di vista funzionali, si comportano come i neuroni reali.
Sebbene sia improbabile che il cervello utilizzi una procedura di apprendimento simile alla retropropagazione, ciò non è molto importante, essendo tale procedura solo uno dei tanti metodi per minimizzare l'errore. Tuttavia, un elemento essenziale di ogni procedura di apprendimento supervisionato è il fatto che la rappresentazione interna, che riflette alcune caratteristiche proprie del compito computazionale, si sviluppi autonomamente, senza bisogno di essere progettata in modo prefissato. La procedura di apprendimento può così fornire informazioni utili sulla relazione tra compiti computazionali e forme di rappresentazione, fornendo eventuali indizi sulle rappresentazioni interne nel cervello.
Modelli neurali per la rappresentazione a opponenza cromatica
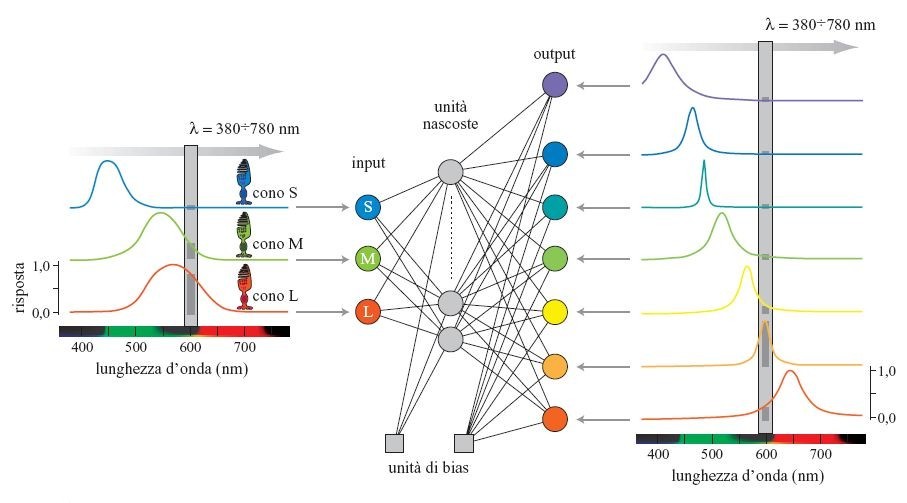
Allo scopo di motivare la presenza di rappresentazioni a opponenza cromatica nel sistema visivo, è stato realizzato un modello neurale a tre strati, il cui compito specifico era quello di discriminare le componenti spettrali dei segnali luminosi (Usui et al., 1994). La rete presenta tre unità di input e sette unità di output, come mostrato in figura (fig. 1). Per mezzo della procedura di retropropagazione, il modello viene addestrato a costruire la corrispondenza tra i segnali di colore tricromatici a banda larga, presenti a livello sensoriale, e i segnali di colore corticali di sette unità cromaticamente selettive. Le unità di input corrispondono alle tre classi di coni nella retina, e le unità di output sono progettate in modo da avere sensibilità spettrali a banda stretta, in accordo con i dati fisiologici sui neuroni selettivi rispetto al colore.
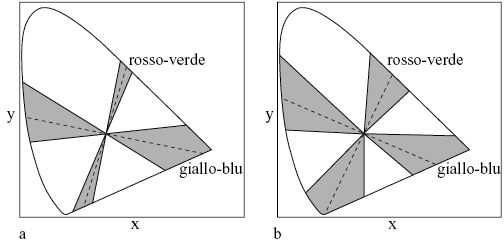
Analizzando le proprietà cromatiche relative ai segnali delle unità nascoste, è stato rilevato (fig. 2) che, in corrispondenza alle linee di sovrapposizione cromatica, sorgono forme caratteristiche di coppie a contrasto cromatico dei tipi rosso-verde e giallo-blu, in analogia a quanto avviene nei neuroni cromatici trovati nel sito LGN del macaco (Derrington et al., 1984). Analizzando la ridondanza delle attività fra unità nascoste, si è anche trovato che le proprietà di risposta spettrale dei modelli che contengono due unità di questo tipo forniscono la rappresentazione (non correlata) più efficiente. Così, il modello con due unità nascoste ha proprietà ottimali per il compito di discriminazione cromatica. È da sottolineare il fatto che la rappresentazione a contrasto cromatico nello strato nascosto si è auto organizzata nella rete per realizzare il desiderato compito elaborativo. In relazione a questo lavoro, G. Buchsbaum e A. Gottschalk (1983) hanno ottenuto risultati simili attraverso una trasformazione Karhunen-Lowe (o analisi delle componenti principali), applicata a triplette di risposte dei coni.
Ricostruzione del solido di Munsell tramite l'elaborazione dei dati fisici di colore
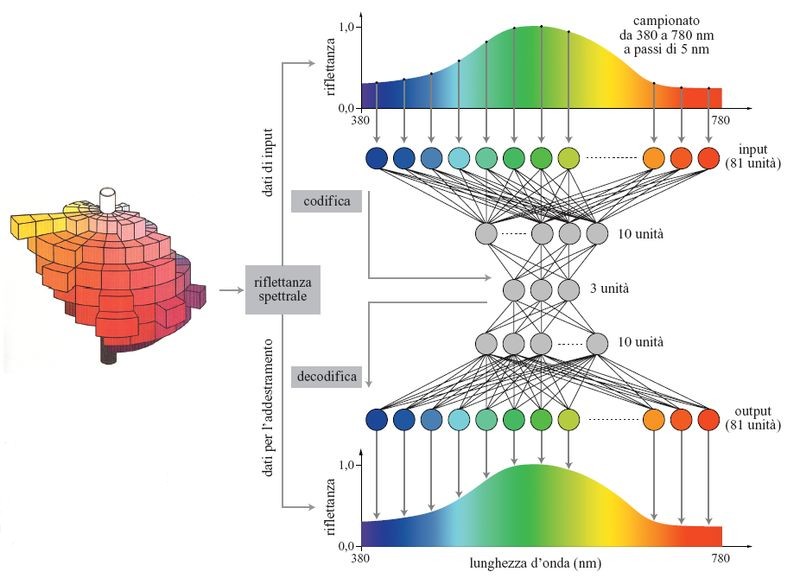
Il colore di un oggetto dipende in modo univoco dalla riflettanza della sua superficie. Le riflettanze superficiali che producono colori discriminabili possono essere rappresentate nel percorso visivo. Per esaminare la rappresentazione efficace delle riflettanze superficiali, è stata generata una cosiddetta corrispondenza identità (che produce un output identico all'input) della riflettanza spettrale superficiale di un solido colorato di Munsell, costituito da 1569 elementi colorati, usando una particolare rete neurale a cinque strati, a forma di calice (Usui et al., 1992). La rete, mostrata in figura (fig. 3), ha un numero di unità nascoste nello strato centrale (il terzo) minore rispetto a quello degli strati di input e di output. Essa è costretta perciò a codificare in modo efficiente l'insieme di dati assegnato, nello strato centrale.
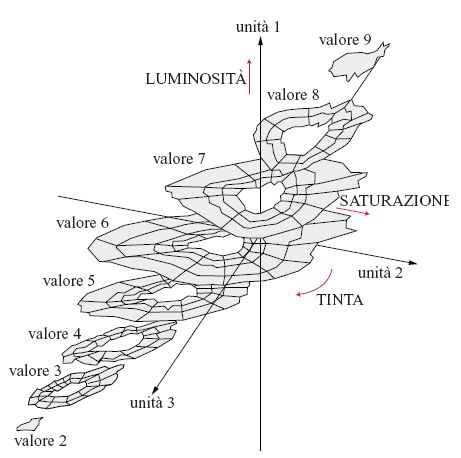
Dopo l'apprendimento, sono stati analizzati gli schemi di attività delle unità nello strato centrale nei casi in cui questo conteneva 2, 3 o 4 unità. Nel caso di 3 unità, una di esse rappresentava il valore di luminosità, e le altre due rappresentavano i due assi di cromaticità. Il solido colorato di Munsell veniva così ricostruito, a partire dai dati fisici, come mostrato in figura (fig. 4). La rete con quattro unità produceva una rappresentazione interna ridondante, mentre la rete con due unità non riusciva a discriminare efficacemente alcune combinazioni di colore. Questi risultati mostrano che le curve di riflettanza spettrale superficiale degli elementi colorati di Munsell possono essere compresse in modo non lineare nello spazio a tre dimensioni, e che l' estrazione non lineare di caratteristiche da parte di una rete neurale a cinque strati è un metodo efficace per determinare la rappresentazione interna nel percorso visivo.
Modelli di visione a colori basati su reti neurali ad apprendimento non supervisionato
Criteri ottimali e apprendimento non supervisionato
È stato dimostrato che alcuni dei criteri ottimali sopra menzionati si possono utilizzare in una procedura di apprendimento non supervisionato di tipo hebbiano. H.E. Barlow (1989) ha proposto il principio di minima ridondanza per una codifica di segnali di input che renda semplice il formarsi di associazioni nuove. Un modo per trovare una codifica di minima ridondanza consiste nel decorrelare il segnale di input. Barlow e P. Foldiak (1989) hanno dimostrato che una rete con connessioni laterali 'retroattive' riesce a trovare una codifica adatta a tale decorrelazione, utilizzando un apprendimento antihebbiano. E. Oja (1989) ha proposto la regola di apprendimento hebbiano per estrarre i vettori di componenti principali della distribuzione degli input. È degno di nota il fatto che l'analisi delle componenti principali è strettamente legata al principio lnfomax (Linsker, 1988; v. anche il saggio di J. Cowan, Storia dei concetti e delle tecniche nella ricerca sulle reti neurali). La preservazione massima di informazione si ottiene attraverso l'analisi delle componenti principali nel caso di un input gaussiano in presenza di rumore. A differenza dell'apprendimento supervisionato, l'apprendimento non supervisionato non richiede segnali esterni. Esso può quindi fornire un metodo efficace per studiare le relazioni tra la rappresentazione dell'informazione e le proprietà statistiche dell'ambiente cui gli stimoli appartengono.
Un modello di rete decorrelante per la costanza cromatica
L'aspetto del colore rimane più o meno costante anche quando si guarda un oggetto in un ambiente variabile. Nel campo della visione a colori la ricerca si è principalmente occupata del fenomeno della costanza cromatica, investigando il meccanismo di percezione invariante del colore. In questo paragrafo descriveremo un modello di rete neurale usata a tale scopo (Usui et al., 1992), che effettua la decorrelazione della tripletta di risposte dei coni a oggetti colorati.
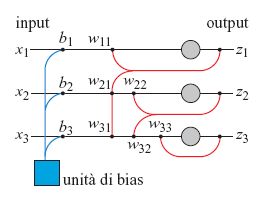
La struttura del modello proposto e la regola di apprendimento sono leggermente diverse rispetto al modello originale (Barlow e Foldiak, 1989). La rete ha tre unità di bias con connessioni modificabili b;, e unità di output con connessioni asimmetriche retro attive wij' come mostrato in figura (fig. 5). Ogni input esterno xi viene trasformato in un segnale interno y;=x;+b;, che viene inviato all'unità di output. Ogni output della rete, Zi, è determinato dal segnale interno y; e dalla somma pesata degli output delle altre unità nel modo seguente:
formula. [1]
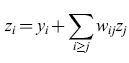
In forma matriciale:
z= (x+b) +wz, [2]
in cui w è una matrice triangolare inferiore (in cui, cioè, sono uguali a zero tutti gli elementi al di sopra della diagonale principale). Dopo il transiente iniziale, possiamo scrivere l' output della rete come
z=(I-W)-ly, [3]
in cui I è la matrice identità. Tutte le connessioni vengono modificate secondo la regola antihebbiana. L'input esterno viene trasformato in un segnale a media nulla fissando le connessioni di bias b; al valore medio di ciascun segnale di input. La regola di apprendimento per le connessioni di bias è:
∆bi = -αYi, [4]
in cui α è il tasso di apprendimento. Le connessioni di retro azione wij vengono modificate in modo da decorrelare le uscite della rete:
formula. [5]
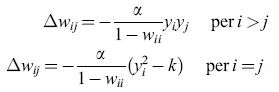
Si noti che la regola di modificazione per le connessioni in autoretroazione wii contiene una costante k che controlla il guadagno dell'output della rete, così che l'intensità di ogni output tende a k durante il processo di apprendimento. Per la fase di apprendimento, è stata usata la tripletta cromatica rosso, verde, blu per le risposte dei coni, illuminando il solido colorato di Munsell con una sorgente di luce bianca (luce diurna) e con due fonti di luce colorata (luce giallastra a incandescenza e luce bluastra a fluorescenza ). lnizialmente i pesi di tutte le connessioni della rete vengono posti uguali azero, e la fase di addestramento ha inizio con i dati relativi alla luce bianca diurna. Terminata tale fase, i valori dei pesi raggiunti vengono assunti come condizioni iniziali per due nuove fasi di apprendimento, rispettivamente con luce a incandescenza e con luce a fluorescenza, ottenendo così tre reti neurali, ciascuna ottimizzata per un tipo di illuminazione.
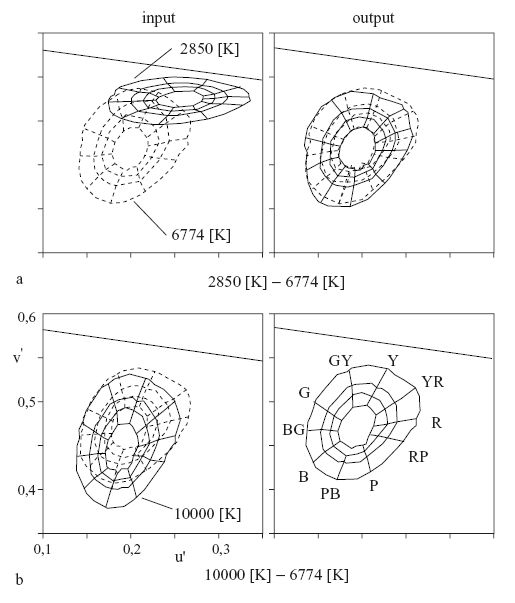
Per valutare quantitativamente l'invarianza della rappresentazione cromatica nel segnale di output della rete, vengono valutate le differenze di colore negli spazi di input e di output della rete, tra le coppie di illuminazione luce diurna-luce a incandescenza e luce diurna-luce a fluorescenza. La figura (fig. 6) mostra gli input e gli output in un piano di valore costante per le due coppie. Ogni griglia nella figura mostra il luogo dei punti di uguale tonalità e saturazione cromatica. Mentre la distribuzione degli input dipende fortemente dal colore della luce illuminante (le distribuzioni per la luce a incandescenza e per quella a fluorescenza si spostano rispetto al caso di luce diurna, fino a diventare rispettivamente giallastra e bluastra), in output si osservano distribuzioni quasi invariate, malgrado il cambiamento della luce illuminante. Pertanto, la rete neurale presenta la proprietà di costanza cromatica, decorrelando le risposte dei coni che dipendono fortemente dal colore della luce illuminante.
Autoorganizzazione dei campi cromatici recettivi in base al principio Infomax
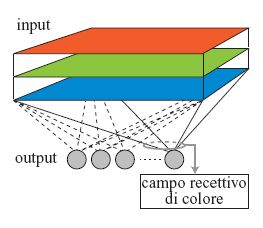
Diversi autori hanno dimostrato che le caratteristiche spaziali dei campi recettivi auto organizzati dipendono fortemente dalle correlazioni spaziali tra i pixel delle immagini usate per l'apprendimento. In questo paragrafo, sarà mostrato come le caratteristiche dei campi recettivi di colore si pongano in relazione con il funzionamento del sistema visivo attraverso un processo di apprendimento non supervisionato basato sul principio Infomax (Usui et al., 1993). Si consideri un modello di rete lineare a due strati (fig. 7), in cui lo strato di input ha tre piani di colore che corrispondono alle matrici bidimensionali dei coni rossi, verdi e blu. Ogni unità di output è connessa con tutti i coni dello strato di input. La risposta della k-esima cellula di output, Yk, è la combinazione lineare delle risposte delle unità di input:
formula [6]
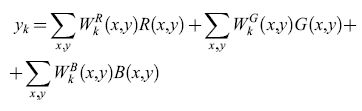
in cui R(xy), G(x,y), B(x,y) sono le risposte di ogni cono in posizione (x,y), e W:(x,y), Wff(x,y), W:(x,y) indicano i pesi delle connessioni assegnate ai coni, che corrispondono al campo recettivo di colore della k-esima unità di output.
Se la distribuzione degli input è gaussiana, e a ogni input viene aggiunto un rumore gaussiano con varianza costante, l'informazione mutua tra input e output viene massimizzata quando i pesi sinaptici corrispondono al vettore principale, nel senso dell'analisi delle componenti principali. Utilizzando la procedura hebbiana generalizzata (Sanger, 1989) per l'aggiornamento delle intensità delle connessioni, la rete converge verso un insieme di pesi sinaptici pari, al limite, proprio ai vettori principali in ordine decrescente di autovalori. Come dati di apprendimento sono usate immagini colorate casuali in presenza di rumore, tali che i campi recettivi cromatici si formino in base a una correlazione non solo spaziale, ma anche cromatica tra i tre tipi di coni: infatti i campi recettivi di colore crescono in funzione della correlazione spaziale e cromatica tra i tre tipi di coni. La statistica delle immagini è definita usando la correlazione tra le risposte di due coni situati in posizioni diverse. La correlazione spazio-cromatica viene descritta moltiplicando la correlazione spaziale per quella cromatica.
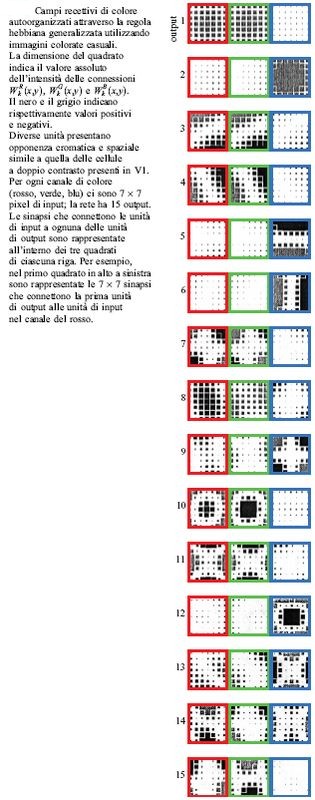
In figura (fig. 8) sono mostrate le intensità sinaptiche autoorganizzate si attraverso la procedura hebbiana generalizzata utilizzando 2500 immagini. Le intensità sinaptiche iniziali sono fissate in modo casuale, la dimensione del piano di colore è di 7 x 7 pixel e il numero di unità di output è pari a 15. l numeri in figura esprimono l'ordine (decrescente) degli auto valori, e da ciò si nota che la prima cellula di output preserva la maggior parte dell'informazione sull'input complessivo. Questo risultato mostra che la prima cellula di output è quella maggiormente efficace nel trattamento dell'informazione dell'immagine di input, in quanto essa preserva la maggior quantità di informazione sensibile agli stimoli (e cioè ai segnali) acromatici. D'altra parte, le cellule numero 2 e 8 si possono interpretare come rivelatori di contrasto passa-banda giallo-blu e rosso-verde, mentre le cellule numero 5, 6 e 13 come rivelatori passa-banda, in accordo con quanto osservato per le cellule a doppio contrasto cromatico presenti nei blob dell'area VI.
Conclusioni
In questo saggio abbiamo messo in evidenza che esistono, in linea di principio, tre tipi di rappresentazione del colore nel percorso visivo, e che le trasformazioni tra queste devono tenere conto di una trasmissione efficace dell'informazione cromatica. Abbiamo descritto vari modelli elaborativi di visione a colori, basati su reti neurali con capacità di apprendimento supervisionato e non supervisionato, per esaminare l' opponenza e la costanza cromatica e i campi recettivi cromatici. Si è così mostrato che l'analisi delle rappresentazioni di colore emergenti dalle reti neurali attraverso la procedura di apprendimento può fornire elementi utili per comprendere l'efficienza della rappresentazione cromatica nel percorso visivo. Il metodo di analisi descritto mostra notevoli potenzialità per lo studio futuro dei meccanismi di elaborazione dell' informazione cromatica nei livelli corti cali superiori.
Bibliografia citata
BARLOW, H.B. (1989) Unsupervised learning. Neural Computation, l, 295-311.
BARLOW, H.B., FOLDIAK, P. (1989) Adaptation and decorrelation in the cortex. In The computing neuron, a c. di Durbin R., Miall Ch., Mitchison G., Workingham-Reading, Addison-Wesley Pub. Co., pp. 54-72.
BUCHSBAUM, G., GOTTSCHALK, A. (1983) Trichromacy, opponent colours coding and optimum colour information transmission in the retina. Proc. R. Soc. London B Biol. Sci., 220, 89-113.
DERRINGTON, A.M., KRAUSKOPF, J., LENNIE, P. (1984) Chromatic mechanisms in lateral geniculate nucleus of macaque. J. Physiol., 357, 241-265.
KOMATSU, H., IDEURA, Y., KAJI, S., Y AMANE, S. (1992) Color selectivity of neurons in the inferior temporal cortex of the awake macaque monkey. J. Neurosci., 12, 408-424.
LENNIE, P., D'ZMURA, M. (1988) Mechanisms of color vision. CRC Crit. Rev. Neurobiol., 3, 333-400.
LENNIE, P., SCLAR, G., KRAUSKOPF, J. (1990) Chromatic mechanisms in striate cortex of macaque. J. Neurosci. 10, 649-669.
LINSKER, R. (1988) Self-organization in a perceptual network. IEEE Computer, 21, 105-117.
LIVINGSTONE, M.S., HUBEL, D.H. (1984) Anatomy and physiology of a color system in primate primary visual cortex. J. Neurosci., 4, 309-356.
OJA, E. (1989) Neural networks, principal components, and subspaces. Int. J. Neural Systems, l, 61-68.
SANGER, T.D. (1989) Optimal unsupervised leaming in a singlelayer linear feedforward neural network. Neural Network, 2, 459-473.
SCHEIN, S.J., DESIMONE, R. (1990) Spectral properties of V4 neurons in the macaque. J. Neurosci., 10, 3369-3389.
SCHEIN, S.J., MARROCCO, R.T., DE MONASTERIO, F.M. (1982) Is there a high concentration of color-selective cells in area V 4 of monkey visual cortex? J. Neurophysiol., 47, 193-213.
USUl, S., NAKAUCHI, S., MIYAKE, S. (1994) Acquisition of color opponent representation by a three-Iayered neural network model. Biol. Cybern., 72, 35-41.
USUl, S., NAKAUCHI, S., MIYAMOTO, Y. (1992) A neural network model for color constancy based on the minimally redundant color representation. Proc. IJCNN (Beijin), 2, 696-701.
USUl, S., NAKAUCHI, S., NAKANO, M. (1992) Reconstruction of Munsell color space by a five-Iayered neural network. J. Opto Soc. Am. A, 9, 516-520.
USUl, S., NAKAUCHI, S., TAKAHASHI, K. (1993) Self-organization of color receptive fields using random color noise images. Proc. WCNN (Portrand), l, 72-75.
ZEKI, S.M. (1980) The representation of colours in the cerebral cortex. Nature, 284, 412-418.
Bibliografia generale
CHURCHLAND, P.S., SEINOWSKI, T.J. The computational brain. Cambridge, Mass., MIT Press, 1992.
WANDELL, B.A. Foundations oJvision. Sunderland, Sinauer Associates Inc., 1995.
ZEKI, S.A. Vision of the brain. Oxford-Boston, Blackwell Sci. Pub., 1993.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


