Selezióne naturale
Enciclopedia on line
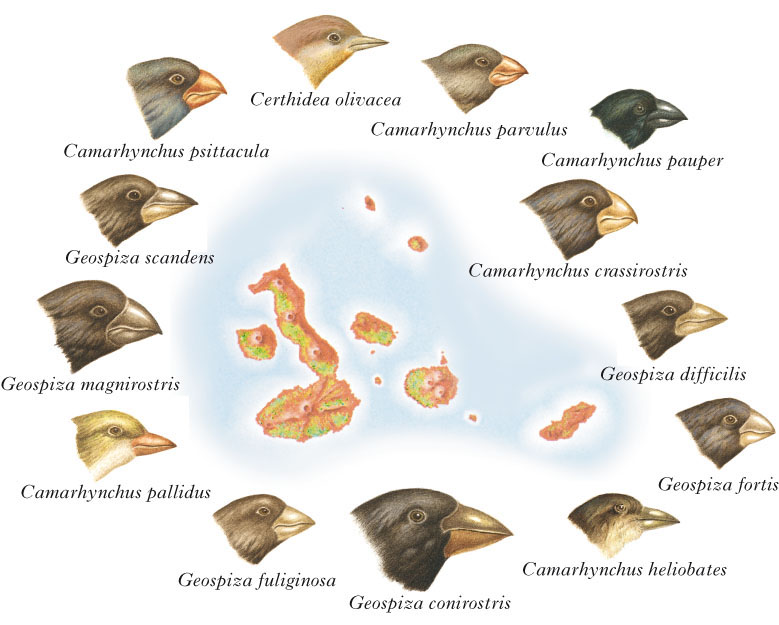
selezióne naturale Meccanismo evolutivo proposto dal naturalista britannico C.R. Darwin nell'ambito della sua teoria dell'evoluzione ed esposto nel libro Sull'origine delle specie per selezione naturale (1859). Darwin, osservando le differenze fra specie affini viventi nelle diverse isole dell'Arcipelago delle Galápagos, si convinse che la lenta modificazione delle specie, la loro evoluzione, quindi, era dovuta principalmente alla s.n.: sopravvivono e si riproducono, cioè, gli individui dotati di caratteristiche più vantaggiose nella lotta per l'esistenza (in sostanza, meglio adattati all'ambiente). In base alle attuali conoscenze di genetica, la s.n. è oggi interpretata come il fattore casuale che favorisce l'aumento di determinate frequenze alleliche nei geni di una popolazione (→ neodarwinismo).
Variabilità genetica e sopravvivenza
Nelle popolazioni naturali il numero dei figli generati è sempre molto elevato, tuttavia solamente una parte di questi giunge a completare lo sviluppo e a riprodursi. Se infatti intervengono mutamenti nell'ambiente, riescono a sopravvivere soltanto gli individui più adatti a tali mutate condizioni, che sono tanti più quanto maggiore è l'assortimento degli alleli, ossia la variabilità genetica nella popolazione. Il processo di s.n. può agire solo in presenza di variabilità genetica, cioè se nella popolazione, in conseguenza di mutazioni, sono presenti almeno due alleli diversi per un determinato gene. In questo modo le frequenze dei fenotipi (cioè delle caratteristiche morfologiche e funzionali degli organismi) che compongono ciascuna popolazione non permangono immutate nel corso delle generazioni, ma si modificano nel tempo; dunque tramite la s.n. si determina l'evoluzione intraspecifica delle popolazioni e, quindi, quella delle specie.
I tre tipi di selezione naturale
A seconda della frequenza con cui si manifestano i diversi fenotipi in una popolazione, si distinguono: una s.n. direzionale, che agisce a favore dei fenotipi corrispondenti a una delle estremità dell'intervallo della curva di variazione fenotipica della popolazione stessa; una s.n. divergente o disruptiva, se sono favoriti gli individui che si trovano in prossimità di entrambi gli estremi; e una s.n. stabilizzante, quando gli individui favoriti sono quelli che presentano un fenotipo che riguarda la media della popolazione.
Selezione sessuale e artificiale
Si ha selezione sessuale quando la s.n. riguarda la scelta di un partner riproduttivo, operata in base alla idoneità dei caratteri sessuali secondari; analogamente, si ha selezione artificiale quando gli accoppiamenti selettivi sono operati dall'uomo in specie animali o vegetali, al fine di ottenere varietà che abbiano caratteristiche particolari, essenzialmente per una migliore utilizzazione economica e produttiva, o a fini estetici.
Focus
La microevoluzione della Biston betularia
Un classico esempio di meccanismo di azione della s.n. è quello descritto per la falena Biston betularia, molto comune in Inghilterra. Prima del 1850, il 99% della popolazione di falene aveva ali di colore grigio maculato di bianco, che consentiva all'individuo di mimetizzarsi perfettamente con la corteccia delle betulle coperte di licheni. Gas tossici prodotti dalle industrie uccisero i licheni e la fuliggine scurì le cortecce, per cui le falene con la colorazione chiara divennero un facile bersaglio per i predatori; le rare farfalle della stessa specie con il fenotipo mutante ad ali scure ottennero invece un grande vantaggio selettivo, potendosi mimetizzare e quindi sottraendosi alla predazione. In pochi anni, si verificò un rapido aumento della frequenza del fenotipo scuro, che divenne prevalente. Più recentemente, miglioramenti delle condizioni ambientali e l'abbattimento dello smog hanno nuovamente favorito la crescita dei licheni e, di conseguenza, il vantaggio selettivo della forma chiara.
Abstract di approfondimento da Evoluzione. Selezione naturale di Saverio Forestiero (Enciclopedia della Scienza e della Tecnica)
Selezione sessuale
Il successo riproduttivo di un individuo non dipende soltanto dalla sua adattabilità ai fattori fisico-chimici e a quelli biologici, interspecifici, rappresentati da competitori, predatori e parassiti, ma è influenzato in grande misura anche dai rapporti, intraspecifici, che ogni adulto intrattiene con i potenziali partner dell’altro sesso. C’è selezione sessuale ogni volta che uno dei due sessi influisce sulla fitness dei caratteri espressi nell’altro individuo attraverso la scelta del partner in vista dell’accoppiamento.
In molti animali gli individui dei due sessi sono facilmente distinguibili sulla base di caratteri somatici extragonadici. Tra questi caratteri sessuali secondari, i più vistosi comprendono l’apparato genitale e i caratteri morfologici extragenitali come, per esempio, la criniera del leone, lo sperone del gallo, il piumaggio vistoso dei maschi di molti uccelli, la presenza di corna nei maschi dei cervidi e in certi coleotteri, le dimensioni corporee. Un forte dimorfismo sessuale può anche interessare i caratteri fisiologici e soprattutto quelli etologici, come tipicamente accade per le molto elaborate e bizzarre pergole di accoppiamento costruite dal maschio degli uccelli giardiniere. Da molto tempo i ricercatori si sono interrogati sull’esistenza nei maschi di eventuali vantaggi collegati al possesso di strutture che tuttavia sembrano ostacolare la loro stessa sopravvivenza. È dimostrato infatti che l’esibizione di livree vistose da parte di molti uccelli possono esporli a una maggiore predazione, sicché qui come in casi analoghi, l’esistenza di un forte dimorfismo sessuale pone un problema esplicativo piuttosto complesso. Perché i maschi sono diversi dalle femmine? C’è un vantaggio nel dimorfismo sessuale, e qual è?
Darwin affronta la questione nel quarto capitolo di On the origin of the species by means of natural selection, (1859) e in tutta la seconda parte di The descent of man, and selection in relation to sex (1871), proponendo due spiegazioni. La prima è che i caratteri sessuali secondarî sarebbero utili ai maschi nella lotta per accedere alle femmine; la seconda è che, siccome le femmine di certi uccelli durante il corteggiamento sembrano essere attratte da caratteri ornamentali come le code vistose e il piumaggio elaborato dei maschi (esempi canonici ne sono rispettivamente la coda del pavone o degli uccelli vedova e le livree degli uccelli del paradiso), questi caratteri sessuali secondarî potrebbero essersi affermati perché attraenti per le femmine. Per questa via Darwin arriva a configurare due modalità di selezione sessuale; quella cosiddetta intrasessuale, attuata attraverso competizione tra maschi per la femmina con cui accoppiarsi, e quella intersessuale attuata dalla femmina quando sceglie tra maschi. Il terzo processo attraverso cui sappiamo che la selezione sessuale agisce è la competizione spermatica, un fenomeno che Darwin conosceva ma cui non diede grande peso. Darwin in effetti ricorre a due principî e fornisce una spiegazione che sembra duplice ma che in realtà spiega solo una metà (la selezione intrasessuale) del fenomeno, lasciando inesplicata la scelta operata dalla femmina (la selezione intersessuale).
Alfred R. Wallace, coautore della teoria della selezione naturale, accettò l’idea di competizione tra maschi per l’accesso alle femmine, ma rifiutò nettamente, giudicandola assurda, l’idea di una femmina capace di scegliere il maschio con cui accoppiarsi e quindi respinse la spiegazione darwiniana dell’evoluzione degli ornamenti dei maschi. L’interesse per i varî aspetti della sessualità in senso lato (la sua origine determinazione ed evoluzione, il dimorfismo sessuale, i sistemi nuziali, la selezione sessuale, il rapporto numerico tra maschi e femmine di una popolazione – sex ratio, le cure parentali), è sempre rimasto vivo e la ricerca è ripartita ogni volta dalla posizione di Darwin. La doppia ipotesi darwiniana fu affrontata per la prima volta sul piano teorico da Ronald Fisher che si propose di superare l’empasse in cui veniva a trovarsi la teoria della selezione naturale impossibilitata a spiegare le ragioni del dimorfismo sessuale e segnatamente i tratti appariscenti di certi maschi come per esempio gli ornamenti degli uccelli del paradiso oppure i giardini e i pergolati degli uccelli giardinieri. D’altra parte se era fuori discussione l’esistenza di una scelta da parte delle femmine, non era invece altrettanto palese quale potesse essere per una femmina il beneficio derivante da questa sua capacità di scegliere un maschio vistoso con cui accoppiarsi.
Nel suo The genetical theory of natural selection del 1930, Fisher elaborò un modello unitario (conosciuto come runaway hypothesis) dell’evoluzione dei caratteri maschili appariscenti e della preferenza femminile dove i tratti maschili e quelli femminili sono fatti coevolvere fino a raggiungere un equilibrio. Fisher interpretò correttamente la preferenza delle femmine per gli ornamenti vistosi come un adattamento raggiunto attraverso l’azione della selezione naturale. La sua ipotesi era che di per sé non vi fossero differenze biologiche rilevanti tra maschi appariscenti e maschi non appariscenti; giudicò costoso esibirsi e quindi giudicò svantaggiati in termini di selezione naturale i maschi portatori di ornamenti costosi e che perciò sprecano risorse, ma convenne che le femmine rispondono molto di più al corteggiamento di questi maschi esibitori di segnali costosi piuttosto che al corteggiamento dei maschi poco appariscenti. In sostanza, secondo Fisher, le femmine tendono a considerare più attraenti i maschi riccamente ornati, i quali perciò da questa tendenza ricaveranno un premio riproduttivo. Secondo questo modello, visto che attraggono un maggiore numero di femmine, solo i maschi sono avvantaggiati; le femmine sono svantaggiate quando generano figli appariscenti, D’altronde, però, solo questo tipo di prole attirerà le femmine e quindi la vistosità dei maschi si perpetuerà nella popolazione attraverso le generazioni.
Una diversa spiegazione della presenza di ornamenti vistosi e stravaganti nei maschi di alcuni uccelli venne data negli anni Settanta da Atmotz Zahavi che ipotizzò che caratteristiche come la coda del pavone rappresentino dei veri e propri handicap per il portatore. Questi handicap, d’altra parte, proprio in quanto tali agirebbero come avvertimenti ‘onesti’ della vitalità del maschio. Osservando la capacità di un maschio di tollerare il proprio handicap, la femmina potrebbe perciò scegliere i maschi migliori con cui accoppiarsi. Ecco allora spiegato come i geni codificanti per questi fenotipi si diffonderebbero nella popolazione. Secondo Zahavi le femmine valuterebbero la qualità dei maschi sfruttando le informazioni rappresentate dall’esibizione di segnali molto costosi come sono le strutture ornamentali corporee. La principale obiezione al modello di Zahavi riguarda la dinamica temporale della variazione fenotipica (e quindi della variazione genetica sottostante). L’ipotesi di Zahavi prevede che col tempo le differenze tra maschi debbano ridursi fino a scomparire per controselezione delle varianti con minore fitness; ma questa previsione è contraddetta dai fatti.
Una soluzione a questo problema venne avanzata nel 1982 da William Hamilton e Marlene Zuk che proposero di interpretare caratteri appariscenti ed esibizioni maschili come handicap rilevatori della resistenza, controllata geneticamente, all’infezione da parte di parassiti. Dimostrare la correttezza dell’ipotesi di Hamilton e Zuk è difficile, mentre è facile obiettare che una femmina potrebbe evitare di accoppiarsi con un maschio malato (e quindi senza ornamenti) semplicemente per evitare di contrarre essa stessa i parassiti. Mentre nel modello di Zahavi tutti i maschi investono in eguale misura nell’handicap, ma solo i più vitali possono sostenere il costo di un tale investimento (l’handicap è dipendente dallo stato di benessere dell’animale), nel modello di Hamilton e Zuk solo i maschi migliori sono portatori di handicap (l’handicap è una spia del vigore dell’animale).
A partire dagli anni Settanta del secolo scorso, le osservazioni e gli studi sperimentali sull’evoluzione dei caratteri sessuali secondari hanno ricevuto nuovo impulso dalla messa a punto di tecniche e di metodi innovativi che hanno permesso di stimare parametri (come, per es., il numero di copule, il loro costo metabolico, la quantità di figli generati, ecc.) influenti sulla fitness di un individuo, hanno consentito di accertare la paternità (mediante il test del DNA fingerprint) e di valutare fattori e modelli comportamentali coinvolti nell’evoluzione della sessualità; in particolare quelli relativi alle colorazioni maschili, alla scelta del partner, alle cure parentali. Tra le domande più frequenti c’è quella se gli animali scelgano o meno il partner in base al fatto che esso possegga caratteri appariscenti. In alcuni casi è quello che avviene. Nel 1982, ipotizzando che nella Vedova codalunga, la specie africana poliginica Euplectes progne, la selezione sessuale favorisse l’evoluzione di code lunghe, l’etologo Malte Andersson manipolò la lunghezza della coda accorciandola ad alcuni esemplari e tagliandola e poi rincollandola (perciò senza alterarne la lunghezza) ad altri. La Vedova codalunga è un uccello dalle dimensioni di un passero caratterizzato da un marcato dimorfismo sessuale: le femmine hanno una coda di circa 7 cm, una lunghezza giudicata pressoché ottimale per il volo, mentre la coda dei maschi può raggiungere il mezzo metro. In natura, i maschi difendono un’area dove si esibiscono in complessi rituali di corteggiamento. Lo sperimentatore dimostrò che la lunghezza della coda non aveva alcun peso nella competizione tra maschi durante la fase di difesa delle arene di corteggiamento, mentre ne aveva nella loro capacità di suscitare l’interesse delle femmine. I maschi con coda accorciata artificialmente attraevano infatti meno femmine di quelli con coda illesa o forniti di code tagliate e rincollate. I maschi con le code allungate avevano arene con un maggiore numero di nidi rispetto a quelle degli altri maschi.
Diversamente dal punto di vista tradizionale che giudicava corteggiamento e accoppiamento come processi eminentemente cooperativi, oggi ha sempre più spazio l’ipotesi dell’esistenza di un vero e proprio conflitto di interesse tra maschio e femmina. Questo in realtà non deve stupire perché, anche se è vero che entrambi i partner sono beneficiati dalla riproduzione, esiste tuttavia una fondamentale asimmetria di ruoli generata dal differente investimento parentale (allocazione di risorse necessarie a nutrire e allevare la prole) che controlla la fitness maschile e quella femminile. La principale differenza tra maschio e femmina risiede nella differente dimensione dei rispettivi gameti (stato di anisogamia): le femmine producono gameti grandi immobili e ricchi di sostanze nutritive, mentre i maschi producono gameti piccoli e solitamente molto mobili (lo spermatozoo comprende una testa contenente DNA e una coda dotata di un flagello propulsore). La situazione è perciò chiaramente distinta: i maschi aumentano la propria fecondità in maniera direttamente proporzionale al numero di accoppiamenti effettuati (vi sarà dunque una selezione sui maschi a favore delle caratteristiche che aumentano la probabilità di essere scelti per l’accoppiamento), mentre le femmine investono nella prole una maggiore quantità di risorse sia nella produzione dei gameti che nelle cure parentali (e perciò la selezione massimizzerà la loro capacità di scegliere i maschi ‘migliori’ insieme a qualunque attributo in grado di favorire l’allevamento della prole). Per questo motivo i maschi sono per loro natura etologicamente ‘focosi’, pronti al combattimento specialmente nel periodo in cui le femmine sono sessualmente ricettive, mentre la diversa condizione delle femmine le rende particolarmente accorte nella scelta del maschio. In molte specie di uccelli e insetti la ritrosia delle femmine viene vinta attraverso un dono nuziale sotto forma di cibo che viene consumato durante il corteggiamento o durante la copula. Anche lo sforzo di accoppiamento (che con l’investimento parentale, rappresenta la seconda componente dello sforzo riproduttivo) differisce tra maschio e femmina. L’investimento necessario per procurarsi il partner è maggiore nel maschio che nella femmina. Complessivamente la selezione sessuale è più intensa nelle specie con sistema nuziale poligamico, promiscuo (cioè un misto di poliginia – il maschio si accoppia con molte femmine e ciascuna femmina si accoppia con un solo maschio – e poliandria – la femmina si accoppia con molti maschi), che nelle specie monogame in cui si stabilisce un legame di coppia e spesso entrambi i genitori concorrono alla cura della prole.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


