Navigazione animale
Enciclopedia Italiana - VII Appendice (2007)
Navigazione animale
I metodi e gli strumenti della navigazione
di Floriano Papi
L'evoluzione naturale favorisce negli esseri viventi l'acquisizione della capacità di spostarsi in maniera efficiente da un luogo all'altro per trovare le migliori condizioni possibili in termini di disponibilità di fonti di cibo e di riproduzione. La navigazione è quindi quell'insieme di capacità che permettono a un animale di raggiungere una meta definita e circoscritta nello spazio, anche se relativamente lontana, quando questa non è già individuabile (cioè semplicemente percepibile) alla partenza mediante un contatto sensoriale diretto. In questo settore di studi la terminologia tecnica è mutuata dall'esperienza umana, cosicché si usano termini nautici come pilotaggio, bussola, mappa e così via; in particolare, in etologia il termine inglese homing (tornare a casa) sta a indicare gli spostamenti dell'animale verso un luogo ben conosciuto, come appunto può essere il nido.
Il metodo storicamente più usato dai naturalisti in questo tipo di studi, è il sistema cattura-contrassegno-ricattura: l'animale viene catturato alla partenza di un viaggio migratorio, viene contrassegnato con un anello, una targhetta o gli viene applicato un transponder magnetico e poi viene rilasciato. In base al luogo in cui avviene la successiva ricattura è possibile inferire il luogo di arrivo o di sosta, ma non la rotta precisa seguita dall'animale. Tale metodo è ovviamente poco efficiente a causa della bassa percentuale di animali ricatturati. I metodi utilizzati tra la fine del 20° e l'inizio del 21° sec. si basano invece sulla telemetria (anche satellitare). Sull'animale catturato viene impiantato un piccolo apparecchio registratore, che per tutta la durata del viaggio rileva sia dati ambientali (temperatura, pressione, luminosità ecc.) sia dati fisiologici dell'animale (attività, frequenza cardiaca, temperatura corporea). L'apparecchio comunica periodicamente questi dati a un sistema GPS (Global Positioning System), che quindi rileva anche istantaneamente longitudine, latitudine e altitudine a cui si trova l'animale.
Altri metodi di ricerca molto fecondi prevedono la realizzazione di esperimenti di laboratorio su animali in cattività. In questi esperimenti, solitamente, si sfrutta la tendenza dell'animale a fuggire verso direzioni determinate, partendo da arene o gabbie circolari, oppure si studiano le modalità con cui l'animale, tipicamente un ratto, esplora un apparato sperimentale a forma di labirinto per ritrovare porzioni di cibo o un riparo. Il laboratorio, evidentemente, comporta per lo sperimentatore il vantaggio di poter manipolare le informazioni spaziali che l'animale utilizza per determinare la direzione di movimento, come, per es., il campo magnetico, le configurazioni stellari o la forma del labirinto.
La maggior parte degli studi etologici sulla navigazione sono stati condotti su uccelli di varie specie, che in natura coprono grandi distanze per trovare il cibo o luoghi adatti per la riproduzione. Tuttavia, molte altre specie animali sono in grado sia di seguire linee guida del paesaggio, come fiumi, valli o coste, sia di mantenere una rotta precisa in un ambiente uniforme, per es. nuotando in mare aperto o sorvolando deserti. Si è visto così che le capacità di orientamento degli animali sono talmente efficienti da consentire loro anche di reagire, compensandoli adeguatamente, a dirottamenti accidentali dovuti, per es., a tempeste, oppure a dislocamenti intenzionali praticati dall'uomo, come nel caso degli studi sui piccioni viaggiatori.
Questi metodi di studio hanno permesso di descrivere con precisione un certo numero di processi comportamentali grazie ai quali un animale può muoversi efficacemente nello spazio.
Gli strumenti di navigazione: bussole e mappe
La navigazione consiste essenzialmente in un processo comportamentale attivo, da parte dell'individuo, reso possibile dalla elaborazione di informazioni di varia origine: da un lato, informazioni sensoriali, prodotte dalle bussole biologiche, dall'altro, informazioni cognitive, incluse in rappresentazioni mentali di aree geografiche più o meno estese chiamate appunto mappe cognitive.
Gli strumenti di navigazione: bussole e mappe
La navigazione consiste in un processo comportamentale attivo, da parte dell'individuo, reso possibile dalla elaborazione di informazioni di varia origine: da un lato, informazioni sensoriali, prodotte dalle bussole biologiche, dall'altro, informazioni cognitive, incluse in rappresentazioni mentali di aree geografiche più o meno estese chiamate appunto mappe cognitive.
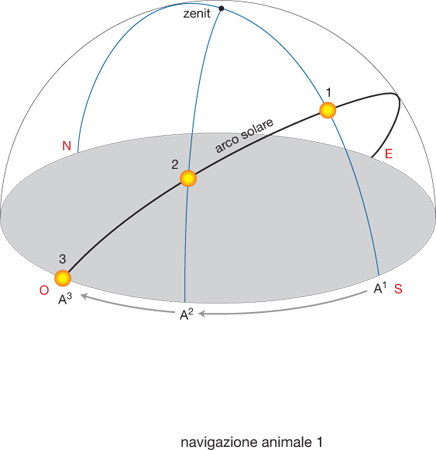
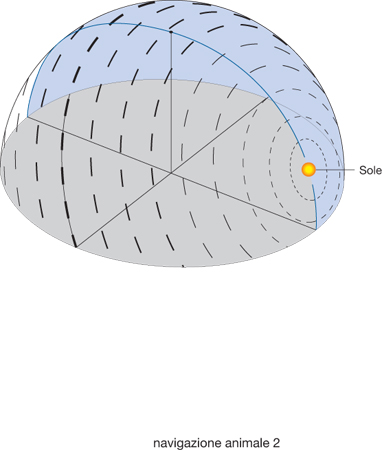
Le bussole biologiche. - Per quanto riguarda le bussole biologiche, la ricerca ha dimostrato che ne esistono diversi tipi. La bussola solare, per es., consente all'animale di orientarsi in base alla posizione del Sole e al suo movimento apparente nel cielo. Questo meccanismo di orientamento si basa sul movimento del Sole lungo l'arco solare, come illustrato nella fig. 1, dove viene esemplificato un arco solare a 45° di latitudine N, in corrispondenza degli equinozi del 21 marzo e del 23 settembre. In figura sono rappresentate le posizioni del Sole a mezzogiorno (1), a metà pomeriggio (2) e al tramonto (3) e i corrispondenti azimut (A1, A2, A3), dove A1 corrisponde al S e A3 all'O. Si noti che il tempo impiegato dall'azimut del Sole per passare da A1 ad A2 è uguale a quello da A2 ad A3 e che la velocità di cui l'animale deve tener conto, pertanto, è diversa. Sono tracciati anche il meridiano celeste che passa per A1-1-zenit e la sua prosecuzione oltre lo zenit, ossia il suo antimeridiano, e il meridiano passante per A2-2-zenit. Le frecce in basso nella fig. 1 indicano la direzione dello spostamento dell'azimut. Prima di poter usare appropriatamente la bussola solare, però, l'animale deve apprendere a riconoscere la posizione del Sole nelle varie ore del giorno rispetto alle direzioni cardinali e nelle diverse stagioni. Successivamente, per riconoscere, assumere e mantenere una data direzione geografica, deve solo conoscere l'ora del giorno per determinare l'angolo di orientamento corretto rispetto al Sole. Questa informazione gli viene fornita da un ritmo biologico circadiano, l'orologio biologico, sincronizzato in base all'alternanza giorno-notte. Ancora poco conosciuto è il processo che permette agli animali di tenere conto della diversa velocità con cui varia l'azimut solare (ossia la sua posizione sul piano dell'orizzonte) a seconda delldel giorno, della stagione e della latitudine. Un'ulteriore fonte di calibrazione della bussola solare rispetto ai punti cardinali è la capacità di alcune specie (come uccelli, api e formiche) di percepire la variazione della percentuale di luce polarizzata celeste e la composizione spettrale della luce. Infatti, l'orientamento del piano di polarizzazione e la percentuale di polarizzazione variano regolarmente e simmetricamente nel cielo, a partire dalla posizione del Sole, il quale determina due emicalotte di luce polarizzata simmetriche separate dal meridiano e dall'antimeridiano solare, e quindi dall'azimut del Sole. Questo fatto è esemplificato nella fig. 2, che offre una rappresentazione schematica della polarizzazione della luce nella volta celeste; la direzione di polarizzazione è fornita dalla direzione delle barre che sono disposte concentricamente rispetto al Sole, rappresentato a un'altezza di 24° sull'orizzonte. Lo spessore delle barre indica la percentuale di luce polarizzata rispetto al totale della luce. La linea passante per il Sole rappresenta il meridiano e l'antimeridiano solare.
L'esistenza di una bussola stellare è stata dimostrata con esperimenti condotti su alcuni Passeriformi (come la specie nordamericana Passerina cyanea) mantenuti in cattività in un planetario, dove, facendo ruotare a piacere la volta stellata, è possibile influenzare la direzione dei tentativi di fuga degli uccelli tenuti all'interno di gabbie circolari. Questa forma di orientamento è basata sull'analisi da parte dell'animale di complesse configurazioni stellari e non sulla posizione di singoli astri, e viene appresa durante le prime settimane di vita grazie all'attenta osservazione della rotazione di tutte le altre stelle intorno alla stella polare.
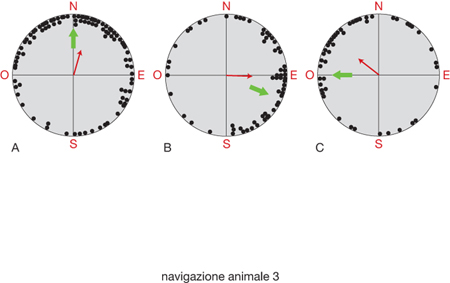
Infine, la bussola magnetica si basa sulla percezione delle variazioni del campo magnetico terrestre (per quanto riguarda polarità, inclinazione e intensità) in funzione delle sue variazioni geografiche. È possibile dimostrare in modo inequivocabile l'esistenza della bussola magnetica mediante esperimenti di laboratorio su Passeriformi, le cui direzioni di fuga possono essere influenzate da manipolazioni del N magnetico. La fig. 3 illustra, per es., il meccanismo di orientamento dei pettirossi (Erithacus rubecula), ingabbiati all'epoca della migrazione primaverile, quando mostrano la tendenza a dirigersi verso N. Nel primo caso (fig. 3 A) gli esperimenti si sono svolti in campo magnetico naturale (la freccia verde indica il N magnetico); nel secondo (fig. 3 B) e nel terzo (fig. 3 C) il N magnetico è stato ruotato rispettivamente in E-S-E e in O, inducendo un parallelo cambiamento nell'orientamento dei pettirossi. Ciascun punto nero indica la direzione media calcolata in base ai tentativi di fuga compiuti da un pettirosso in una notte, la freccia rossa, invece, indica il vettore risultante dalla distribuzione dei singoli punti. Quello che gli uccelli percepiscono effettivamente è l'inclinazione del vettore magnetico rispetto alla superficie terrestre: tutto ciò è possibile perché l'inclinazione del vettore è nulla all'equatore e cresce gradualmente in senso opposto man mano che ci si sposta in direzione dei poli.
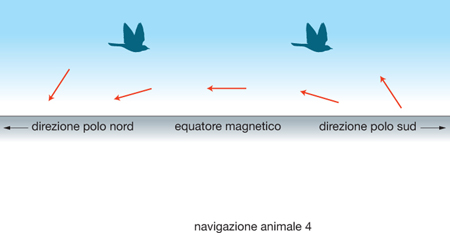
In tal senso, la fig. 4 mostra uno schema della situazione che deve affrontare un uccello che migri dall'emisfero boreale a quello australe basandosi sulla sua bussola magnetica. Il vettore magnetico è indicato dalle frecce: se l'uccello ne percepisse la polarità, saprebbe in ogni momento distinguere il N (indicato dalla testa della freccia) dal S. Essendo invece sensibile solo all'inclinazione del vettore, non può orientarsi dove l'inclinazione è pari a zero. Altrove l'uccello può dedurre, dall'inclinazione del vettore, la direzione del polo magnetico più vicino, che è quella verso cui il vettore è rivolto in basso. La direzione opposta invece è quella dell'equatore magnetico: infatti, nell'emisfero boreale l'estremità del vettore che indica il N punta verso il basso, mentre nell'emisfero australe punta verso l'alto. È ancora un problema irrisolto l'individuazione precisa del recettore sensoriale che consente la percezione del campo magnetico terrestre, ma è noto che negli Elasmobranchi esistono organi elettrorecettori, le ampolle di Lorenzini, in grado di rilevare la presenza e l'orientamento di campi elettrici attorno all'animale.
In molte specie animali, soprattutto negli uccelli migratori, coesistono più tipi di bussola e la navigazione è basata sulla reciproca e continua interazione, calibrazione e sussidiarietà tra questi. Numerosi esperimenti di laboratorio infatti hanno dimostrato come, sia durante le prime settimane di vita sia nel corso di svariate esperienze migratorie, gli uccelli riescano a compensare le imprevedibili variazioni nella disponibilità di un certo indice per l'orientamento (per es., quando la visione diretta del Sole è impedita da spesse nubi o da accorgimenti sperimentali in laboratorio), grazie a un qualsiasi altro indice liberamente disponibile, come, per es., il campo magnetico terrestre o la luce polarizzata. Inoltre, è noto che la bussola magnetica viene costantemente calibrata durante tutto il corso di una migrazione, grazie alla bussola solare.
Le mappe. - Per quanto concerne le mappe cognitive, dal comportamento dell'animale in natura o in laboratorio è possibile indubbiamente inferire l'esistenza di rappresentazioni mentali, immagazzinate nella memoria a lungo termine dell'individuo, che contengono informazioni relative alla posizione reciproca di diversi punti di riferimento in una certa area geografica. Tali rappresentazioni, dette appunto mappe mentali o cognitive, sono apprese mediante ripetute esperienze di navigazione e consentono all'animale di calcolare qualunque percorso possibile a partire da un certo punto di partenza verso un dato punto di arrivo. I punti di riferimento ambientali grazie ai quali vengono elaborate le mappe cognitive sono spesso molteplici e consistono in stimoli di natura visiva, chimica (olfattiva) e acustica. In alcuni casi, come, per es., per i piccioni viaggiatori, la navigazione è possibile anche attraverso aree sconosciute, qualora però esse siano attraversate da gradienti olfattivi le cui variazioni geografiche sono note all'animale (v. oltre).
I meccanismi della navigazione
È possibile classificare le strategie di navigazione esibite dagli animali in base alla fonte delle informazioni necessarie per elaborare la rotta da seguire. Bisogna ancora sottolineare, tuttavia, che molte fra le specie animali possono avvalersi alternativamente o contemporaneamente di più strategie di navigazione, a seconda di quale sia la migliore in determinate condizioni ambientali. Il tipo più semplice di meccanismo di navigazione si basa su una ricerca sistematica, che, in mancanza di migliori indizi per l'orientamento, viene realizzata esplorando a tappeto e con regolarità tutto il territorio circostante al punto di rilascio, in direzioni concentriche che si allontanano gradualmente da esso. Grazie a questa sistematicità, l'animale è in grado di ritrovare il nido (homing). La strategia di orientamento su base genetica avviene grazie alle informazioni codificate nel genoma della specie riguardanti la direzione, il verso e la lunghezza del viaggio da compiere: essa consente anche a quegli individui che non hanno mai visitato prima il luogo di arrivo di giungere a destinazione. Questa strategia è tipica delle specie più semplici come certi insetti, di cui la farfalla monarca (Danaus plexippus) è un esempio lampante: gli adulti di questa specie nordamericana sono in grado di percorrere almeno 3000 km per raggiungere le residenze invernali, anche se non le hanno mai visitate prima. La strategia di orientamento su tracce precedenti, invece, si basa sul fatto che durante il viaggio di andata l'animale stesso deposita delle tracce fisiche (di solito olfattive) che esso seguirà all'indietro durante il viaggio di ritorno. Queste due ultime strategie, ovviamente, rivestono scarso interesse ai fini dello studio dei processi cognitivi negli animali.
Al contrario, sebbene partano anche questi dal presupposto che si sia svolto un viaggio di andata, esistono altri metodi di orientamento che invece si affidano pesantemente alle abilità di apprendimento dell'animale: è questo il caso della inversione del percorso, vale a dire quando l'animale ripercorre in sequenza inversa la lista di stimoli e di punti di riferimento dell'ambiente che ha memorizzato durante il viaggio di andata. Per es., quando i salmoni del Pacifico (Oncorhynchus nerka) migrano dal mare verso i laghi e i fiumi dove sono nati, per riprodursi e poi morire, riescono a compiere numerose scelte corrette mentre risalgono più fiumi confluenti tra loro, nuotando per migliaia di chilometri grazie appunto alla memoria delle tracce olfattive che caratterizzano l'acqua in corrispondenza di ciascun luogo di svolta, di transito e di arrivo.
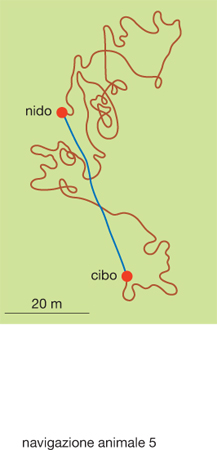
Un secondo esempio di orientamento basato sul viaggio di andata, ben più complesso del precedente, è l'integrazione del percorso (path integration). In questo caso, l'animale tiene conto della direzione e della lunghezza di ogni tratto del percorso di andata ed è in grado in ogni momento di calcolare la propria posizione rispetto al punto di partenza (che di solito è il nido); grazie a ciò, se è necessario, l'animale è capace di far ritorno al nido per la via più breve e diretta a dispetto della tortuosità del viaggio di andata. Ciò è esemplificato nella fig. 5: dopo essere uscita dal nido, l'operaia di una specie di formica deserticola, Cataglyphis albicans, ha esplorato con un tragitto tortuoso (linea rossa) il terreno circostante finché non ha trovato il cibo. Da quel momento è rapidamente ritornata al nido con un percorso rettilineo (linea azzurra). Il percorso illustrato in fig. 5 è stato ricostruito osservando i movimenti della formica all'interno di un reticolato dipinto sul terreno. Le informazioni sensoriali su cui si basa l'integrazione del percorso sono di due tipi: le informazioni allotetiche derivano dalla percezione di stimoli e punti di riferimento ambientali, ossia esterni all'animale; quelle idiotetiche, invece, consistono nella registrazione dei programmi di movimento e delle sensazioni propriocettive elaborate dal sistema nervoso dell'animale. è degno di nota il fatto che, poiché le informazioni idiotetiche sono soggette a un progressivo cumulo di errori in conseguenza della lunghezza del percorso svolto, le informazioni allotetiche vengono utilizzate per la correzione di quelle idiotetiche.
Altra forma di navigazione è il pilotaggio: è quella strategia di orientamento che si basa esclusivamente sul ricordo che l'animale conserva della posizione di un insieme di punti di riferimento (di solito visivi). Esso presuppone, cioè, l'uso da parte dell'animale di una vera e propria mappa cognitiva dell'ambiente elaborata in base all'esperienza precedente. Tuttavia, le forme più complesse di navigazione, definite navigazione vera, consentono all'animale di prescindere almeno in parte da informazioni acquisite durante uno o più viaggi precedenti di andata e gli permettono di raggiungere l'obiettivo anche partendo da zone sconosciute per mezzo di una o più bussole biologiche.
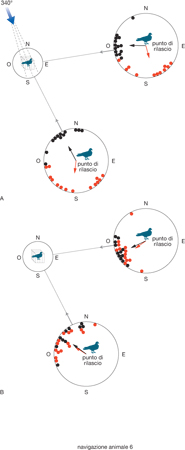
La navigazione vera può essere basata sull'impiego concomitante di una mappa cognitiva dell'ambiente noto e delle bussole biologiche, ma esistono anche casi in cui l'animale può ben orientarsi in luoghi ignoti. È questo il caso del piccione viaggiatore (Columba livia). Questo uccello, infatti, è in grado di orientarsi sia in base alla mappa cognitiva dei luoghi già esplorati in precedenza, sia in base a una mappa olfattiva molto più ampia, acquisita in base all'esperienza degli odori trasportati dai venti; questa mappa si consolida durante le prime settimane di allevamento ed è centrata sulla posizione della colombaia. Nella fig. 6 è mostrato un esempio di mappa olfattiva del piccione viaggiatore, che viene acquisita associando la direzione dei venti e degli odori. Questo fatto può essere dimostrato con metodi sperimentali: due gruppi di piccioni sono stati allevati in gabbie separate, esposte ai venti da ogni direzione. Nella fig. 6 A i piccioni del gruppo sperimentale erano investiti frequentemente anche da un vento artificiale che era prodotto da grossi ventilatori collocati in una direzione vicina a N-N-O (340°), che portava un odore artificiale di mandorle amare ottenuto con benzaldeide. I piccioni sono stati poi trasportati a una cinquantina di chilometri verso E e verso E-S-E e rilasciati. Il risultato è riportato nei due diagrammi (cerchi grandi) della fig. 6, che mostrano le direzioni prese dagli animali dopo essere stati liberati. A una metà di essi era stata fatta odorare benzaldeide prima del rilascio, e questi si sono diretti verso la direzione opposta a quella da cui erano soliti sentire giungere l'odore di mandorle amare; mentre l'altra metà, non esposta alla benzaldeide prima del rilascio, ha puntato verso casa (punti neri). Nella fig. 6 B, i piccioni del gruppo di controllo, che non erano stati esposti al vento odoroso, si sono orientati sempre verso casa, indipendentemente dall'aver fiutato benzaldeide prima del rilascio (punti rossi) o meno (punti neri). Nei diagrammi ogni punto corrisponde alla direzione di un piccione dal punto di rilascio, mentre le frecce interne rappresentano i vettori medi per i singoli gruppi di direzioni. I vettori sono tanto più lunghi, quanto più le direzioni prescelte dai singoli piccioni sono simili tra loro. In base a questi studi, perciò, è chiaro che il piccione, trasportato anche a molti chilometri di distanza o in un luogo non compreso nella mappa cognitiva, è capace di ritornare al luogo di allevamento seguendo il tipico gradiente olfattivo che lo caratterizza. Allo stesso modo, altre specie animali utilizzano come coordinate spaziali le stimolazioni sensoriali prodotte da bussole magnetiche, che possono essere così precise da consentire loro di tracciare la loro posizione (tecnicamente, di fare il punto) addirittura, per es., lungo cospicui tragitti oceanici.
Le basi neurali delle mappe cognitive
di Marco Aversano
Ricerche sperimentali condotte in laboratorio su diverse specie animali (soprattutto ratti) hanno dimostrato che una precisa regione del telencefalo dei mammiferi (il cui omologo negli uccelli è il pallio mediale) è selettivamente deputata all'analisi delle informazioni necessarie per la navigazione: questa regione è l'ippocampo; esso con le sue strutture omologhe si ritrova praticamente in tutti i vertebrati superiori. È noto ormai che in regioni dell'ippocampo, come il campo cellulare CA1 o il subiculum, sono presenti cellule piramidali la cui attività di scarica elettrica è selettivamente dipendente dalla posizione dell'animale in un dato punto noto dell'ambiente (cellule di posizione, place cells), oppure dalla direzione di movimento dell'animale stesso (cellule di direzione, head direction cells).
Questo filone di studi si è dimostrato una preziosa fonte di informazioni comparative valide anche al fine di comprendere il funzionamento della memoria e delle funzioni cerebrali superiori nell'uomo. Infatti, la n. a. può essere considerata un argomento scientifico con risvolti sia di tipo etologico e naturalistico, sia di tipo psicologico e neurobiologico. Se si considera la navigazione come uno dei comportamenti più complessi che gli animali possono esibire, ossia un comportamento che si basa su raffinate capacità di elaborazione sensoriale, di apprendimento e di plasticità della memoria a lungo termine, appare evidente l'analogia con comportamenti umani anch'essi basati sul funzionamento dell'ippocampo, quali, per es., il ragionamento e la memoria. Non solo: diventa anche chiaro che lo studio di modelli animali di laboratorio basati sulla navigazione, come, per es., il comportamento di esplorazione di un labirinto da parte di un ratto, può fornire informazioni preziose sul funzionamento normale e patologico del cervello umano. Tutto ciò è talmente vero che i test comportamentali su animali, finalizzati allo studio delle basi biologiche della memoria, prevedono quasi esclusivamente apparati sperimentali che riproducono situazioni di navigazione o orientamento spaziale all'interno di strutture di laboratorio, come labirinti, arene aperte e così via. Secondo tali ricerche, l'ippocampo sarebbe il fulcro su cui converge un intricato sistema di strutture cerebrali per la memoria e l'apprendimento; esso in effetti riceve afferenze neurali altamente elaborate da numerose aree della corteccia cerebrale, che poi verrebbero sintetizzate in una mappa cognitiva dell'ambiente.
Per gli studiosi la mappa cognitiva rappresenta perciò l'esito finale di un notevole lavoro di integrazione di informazioni provenienti da più sistemi di memoria che sono localizzati all'intero cervello e sono dotate di un significativo grado di complessità computazionale. In questo senso, alcuni autori sostengono che le mappe cognitive elaborate dall'ippocampo non siano altro che un caso specifico di apprendimento (chiamato appunto apprendimento relazionale) di reti di riferimenti tra singoli stimoli o informazioni: tali reti o relazioni tra stimoli possiederebbero proprietà emergenti non prevedibili in base alla semplice somma delle parti che le compongono. Da un punto di vista evoluzionistico, come sottolineano gli studiosi di n. a., è decisamente vantaggiosa la tendenza a costruire mappe cognitive di un dato ambiente mediante molteplici processi neurosensoriali che si attivano in parallelo, tramite i quali l'animale possa sfruttare tutti i possibili indizi sensoriali disponibili. Infatti, questa flessibilità cognitiva permetterà all'animale di ritrovare la strada anche quando l'ambiente conosciuto è cambiato per un qualunque motivo, a causa, per es., di un mutamento oro-geografico. In conseguenza di ciò, la costruzione e l'utilizzo della mappa cognitiva comportano sempre l'interazione e la calibrazione reciproca di singole rappresentazioni mentali basate su un dato indizio sensoriale (come una bussola solare o gli indizi idiotetici propriocettivi) o su una traccia mnestica (come il riconoscimento di una configurazione di elementi oro-geografici) disponibili in un dato ambiente.
Le componenti della mappa cognitiva
Secondo le teorizzazioni più recenti, dunque, la mappa cognitiva emerge dall'interazione di due principali componenti funzionali, distinguibili sia per le sottostrutture ippocampali necessarie per il loro funzionamento, sia per il tipo di dati sensoriali trasferiti nella mappa cognitiva più generale. Le due componenti funzionali agiscono sempre in parallelo per assicurare all'animale un corretto e flessibile orientamento nell'ambiente. La prima di queste due componenti è definita mappa direzionale o di rilevamento (bearing map). Tale rappresentazione mentale è mediata da aree ippocampali come il giro dentato, il campo cellulare CA3 e il sistema fimbria-fornice; questa mappa consente all'animale di calcolare la direzione da seguire in base a stimoli ambientali dotati di un gradiente direzionale preciso (da cui il nome), come, per es., il campo magnetico terrestre o i gradienti olfattivi descritti, o anche in base a caratteristiche geometriche peculiari dell'ambiente di navigazione, come la forma di un labirinto che una cavia da laboratorio può apprendere a esplorare. Grazie alla mappa direzionale, l'animale può navigare anche per lunghissime distanze muovendosi verso (o allontanandosi da) singoli stimoli e gradienti sensoriali distribuiti su ambienti di vasta scala.
La seconda componente è definita mappa visiva (sketch map). Questa rappresentazione mentale contiene il ricordo di configurazioni di percezioni di singoli oggetti o stimoli fisici presenti in un dato ambiente - nonché il ricordo della posizione relativa di ciascuno di essi - e utili per contrassegnare un certo luogo, come boschi, fiumi o montagne, dopo che l'animale ha effettuato un certo numero di viaggi esplorativi e ha esaminato l'ambiente da più punti di vista. Tale rappresentazione viene elaborata all'interno dell'ippocampo grazie alle afferenze corticali e al funzionamento del campo cellulare CA1, e in particolare è resa possibile da ingenti fenomeni di potenziamento a lungo termine (LTP, Long Term Potentiation) nelle cellule ippocampali che sono prodotti dall'attivazione dei recettori post-sinaptici di tipo NMDA (N-metil-D-aspartato), sensibili al neurotrasmettitore glutammato.
L'intera struttura dell'ippocampo è deputata alla integrazione di queste due componenti che agiscono in parallelo, e dunque la mappa cognitiva integrata contiene una matrice incrociata di indizi per l'orientamento, forniti da gradienti fisici nell'ambiente e da tracce di memoria a lungo termine per configurazioni complesse di stimoli che caratterizzano un dato ambiente specifico. Un punto di vista analogo a quello precedente, frutto principalmente di studi sperimentali su roditori, attribuisce all'ippocampo il ruolo di sintesi fra tre dimensioni o processi di elaborazione delle informazioni indispensabili per la navigazione: controllo del movimento, riconoscimento dell'ambiente e informazione percettiva.
La dimensione attinente al controllo del movimento emerge dalla registrazione continua, da parte dell'animale impegnato in un episodio di esplorazione, dell'intera somma di movimenti da e verso un punto di riferimento, di scelte destra o sinistra (o sopra o sotto) di una certa posizione o dalla registrazione della direzione e distanza di movimento da un indizio ambientale rilevante. La dimensione relativa al riconoscimento dell'ambiente implica la percezione e l'immagazzinamento nella memoria a lungo termine di informazioni relative a gruppi di punti di riferimento, traiettorie o assi di movimento possibili in un dato ambiente, a partire da un dato punto di partenza, mediante uno o più specifici punti di orientamento o grazie alla forma generale dei confini dell'ambiente. Infine, la dimensione relativa all'informazione percettiva include la capacità di elaborare, da un punto di vista prima sensoriale e poi cognitivo, la vasta gamma di indizi sensoriali che costruiscono le due dimensioni precedenti; ovvero, la dimensione dell'informazione è la somma delle capacità percettive che consentono a un animale, per es., di calcolare internamente la durata di uno spostamento, oppure di riconoscere un dato oggetto (per es., il paesaggio circostante, la sua tana) o i gradienti visivi e olfattivi dell'ambiente.
Il ruolo dell'ippocampo
È evidente perciò come le concettualizzazioni sulla neurobiologia della n. a., a prescindere dalla maggiore enfasi data a questo o a quell'aspetto teorico, attribuiscano all'ippocampo un ruolo centrale di coordinamento e sintesi dell'intera attività del telencefalo degli animali. Ciò, tra l'altro, comporta notevoli conseguenze sul piano comparativo che spiegano come mai, da un punto di vista evoluzionistico, l'ippocampo susciti un interesse notevole tra gli studiosi. In primo luogo, infatti, l'ippocampo è una struttura che presenta omologie anatomiche in numerose specie di uccelli e mammiferi; in secondo luogo, al di là delle generali differenze anatomo-funzionali nel telencefalo di uccelli e mammiferi, è degno di nota il fatto che le dimensioni dell'ippocampo relativamente all'intero cervello di una specie aumentano in modo prevedibile in base al comportamento spaziale della specie in questione. È stato dimostrato, per es., che negli uccelli e nei roditori possiedono un ippocampo di maggiori dimensioni proprio quelle specie che dipendono fortemente da solide abilità di navigazione per il reperimento delle fonti di cibo, rispetto a specie simili ma caratterizzate da un'ecologia alimentare più stanziale. In breve, nell'ippocampo dei vertebrati superiori (uomo compreso) le rappresentazioni dell'ambiente derivate da singoli e specifici canali sensoriali interagiscono fra loro, formando una rappresentazione sovraordinata che trascende e completa le singole rappresentazioni che la compongono. Da quanto esposto, quindi, è facile dedurre che le prospettive della ricerca futura sulla n. a. riguardano tre aree di studio principali: la descrizione sempre più accurata delle rotte percorse in natura dalle varie specie; l'individuazione più precisa dell'anatomia e della fisiologia delle bussole biologiche; e, infine, l'approfondimento dell'analisi del funzionamento delle strutture cerebrali deputate alla elaborazione e all'immagazzinamento delle informazioni contenute nelle mappe cognitive.
bibliografia
R.R. Baker, The evolutionary ecology of animal migration, London 1978.
J. O'Keefe, L. Nadel, The hippocampus as a cognitive map, Oxford 1978.
Cognitive processes and spatial orientation in animal and man, ed. P. Ellen, C. Thinus-Blanc, Dordrecht 1987.
C.R. Gallistel, The organization of learning, Cambridge (Mass.) 1990.
Animal homing, ed. F. Papi, London-New York 1992.
A.D. Redish, D.S Touretzky, Cognitive maps beyond the hippocampus, in Hippocampus, 1997, 7, 1, pp. 15-35.
H. Eichenbaum, P. Dudchenko, E. Wood et al., The hippocampus, memory and place cells: is it spatial memory or a memory space?, in Neuron, 1999, 23, pp. 209-26.
L.F. Jacobs, F. Schenk, Unpacking the cognitive map: the parallel map theory of hippocampal function, in Psychological review, 2003, 110, 2, pp. 285-315.
R.J. McDonald, N.S. Hong, B.D. Devan, The challenges of understanding mammalian cognition and memory-based behaviours: an interactive learning and memory systems approach, in Neuroscience and biobehavioral reviews, 2004, 28, 7, pp. 719-45.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


