Evoluzione
Enciclopedia del Novecento (1989)
Evoluzione
di Ernst Mayr
SOMMARIO: 1. Introduzione. □ 2. Aspetti dell'evoluzione biologica: a) diversità; b) speciazione; c) discendenza comune; d) l'origine della vita; e) mutamento evolutivo e adattamento; f) la selezione naturale e un processo a due stadi; g) il bersaglio della selezione; h) selezione di sopravvivenza; i) adattamento. □ 3. Limiti all'efficacia della selezione naturale. □ 4. Estinzione. □ 5. Il ruolo del comportamento. □ 6. Selezione sessuale. □ 7. Macroevoluzione. □ 8. Come avviene il mutamento evolutivo. □ 9. La teoria degli equilibri di interpunzione. □ 10. L'origine evolutiva di nuove strutture. □ 11. L'origine di taxa superiori. □ 12. Progresso evolutivo. □ 13. L'evoluzione dell'Uomo. □ 14. Il futuro della ricerca nel campo dell'evoluzione. □ Bibliografia.
1. Introduzione.
Fin dai tempi dei primi filosofi e dei fondatori delle più antiche religioni, l'uomo si è interrogato sull'origine del mondo e sul proprio posto in esso. È vero che l'uomo e il mondo sono soggetti a continui mutamenti, che danno origine a fenomeni sempre nuovi, come suggerisce il concetto di evoluzione? Lamarck (1809) è stato il primo ad aver sviluppato una teoria concreta dell'evoluzione graduale, ma il concetto di evoluzione si è imposto soltanto in seguito alla pubblicazione, avvenuta nel 1859, dell'Origine delle specie di Ch. Darwin. Quello di evoluzione è il concetto che più di ogni altro ha consentito di sviluppare una concezione unitaria in campo biologico: nessun altro concetto scientifico ha contribuito tanto alla comprensione dell'uomo (v. evoluzione: L'evoluzionismo nella cultura del XX secolo). Dato che l'evoluzione è già stata trattata in questa Enciclopedia (v. evoluzione: La moderna teoria dell'evoluzione), questo articolo di aggiornamento si limiterà all'esposizione di argomenti non sufficientemente approfonditi nell'articolo precedente, di nuove scoperte e, infine, di questioni che sono diventate oggetto di discussione solo in anni recenti.
Il termine ‛evoluzione' implica un mutamento graduale, generalmente orientato in una determinata direzione. In particolare l'evoluzione biologica si definisce, più precisamente, come ‛mutamento nella diversità e adattamento di popolazioni di organismi'. Essa riguarda tutti gli aspetti degli organismi: strutture, percorsi di sviluppo, funzioni, specie ed ecosistemi interagenti. Quando Lamarck e Darwin proposero le loro idee, si parlava di ‛teoria' dell'evoluzione; la scienza moderna considera ormai l'evoluzione un fatto altrettanto assodato quanto il fatto che la Terra gira intorno al Sole o che la Terra è sferica e non piatta. Le prove a favore dell'evoluzione sono varie e tutte schiaccianti: tra queste, per esempio, le serie di fossili incluse in strati geologici accuratamente datati mediante precisi metodi radiometrici. L'evoluzione è anche comprovata dal fenomeno dell'ereditarietà, che ci ha insegnato che a ogni generazione si forma un nuovo assortimento di genotipi, e dalle ricerche di biologia molecolare, che hanno consentito la ricostruzione di mutamenti succedutisi nel tempo in determinate molecole, mutamenti cui hanno fatto riscontro quelli di certe caratteristiche strutturali, scoperti da anatomisti e tassonomisti. La certezza che l'evoluzione sia un dato di fatto non è scossa dalle moderne controversie nel campo della biologia evolutiva, controversie che riguardano il come e il perché di specifici problemi evolutivi.
Il problema più dibattuto del darwinismo, fin dall'origine della teoria, è stato quello della causa, ovvero del meccanismo, dell'evoluzione. La teoria di Darwin, quella della selezione naturale, era, nei primi ottanta anni dopo il 1859, la meno popolare fra tutte le teorie alternative. Fra le molte teorie avverse, le più largamente condivise erano tre: il saltazionismo, la teoria dell'autogenesi, od ortogenesi, e il neolamarckismo.
I sostenitori del saltazionismo ritenevano che le specie e le categorie superiori nascessero all'improvviso e che specie, categorie superiori e tutte le più importanti innovazioni evolutive fossero prodotte da macromutazioni in singoli individui.
Secondo la teoria dell'autogenesi, od ortogenesi, l'evoluzione tenderebbe a una perfezione sempre maggiore, guidata da un intrinseco meccanismo finalistico, come, per esempio, il principio omega di Teilhard de Chardin.
Secondo il neolamarckismo, l'evoluzione sarebbe dovuta alla trasmissione ereditaria di caratteri acquisiti ‛modellati' dall'ambiente.
Tutte e tre queste teorie alternative sono state completamente respinte a partire dagli anni quaranta, mentre le posizioni, precedentemente avversate, dei biologi evolutivi hanno ricevuto un ampio consenso, confluendo in quella che J. Huxley ha chiamato la ‛sintesi evolutiva'.
L'evoluzione, secondo la teoria sintetica, è provocata dalla produzione incessante di variazione genetica, attraverso processi casuali, e dal diverso grado di sopravvivenza e di riproduzione dei nuovi individui, geneticamente unici, prodotti da questo processo. Il successo riproduttivo differenziale degli individui è ciò che Darwin ha chiamato ‛selezione naturale'. Bisogna sottolineare il fatto che tutte le recenti controversie sono sorte all'interno della teoria sintetica, mentre tutte le pretese che il darwinismo (o il neodarwinismo) sia da respingere sono basate su incomprensioni, come spiegherò in seguito.
2. Aspetti dell'evoluzione biologica.
I due principali problemi dell'evoluzione sono la spiegazione dell'origine della grande varietà di organismi sulla Terra e la spiegazione degli adattamenti reciproci fra organismi e dell'adattamento degli organismi al mondo in cui vivono. Tutti gli altri aspetti dell'evoluzione rientrano nell'ambito di queste due problematiche.
a) Diversità.
Prima di Darwin, la grande varietà degli organismi veniva spiegata o come prodotto della creazione divina o come dovuta all'origine istantanea, spontanea, di nuovi tipi. Secondo questa concezione saltazionista, le specie si moltiplicherebbero attraverso l'improvvisa comparsa di singoli individui che rappresenterebbero una nuova specie, un nuovo genere, o una nuova categoria superiore. Questo modo di concepire l'origine di nuove specie concordava con l'allora dominante filosofia essenzialista, una filosofia risalente a Platone e, sotto il nome di ‛realismo', largamente condivisa nel Medioevo.
La spiegazione fornita da Darwin per la moltiplicazione delle specie era fondamentalmente diversa. Secondo Darwin le specie erano prodotte attraverso la modificazione graduale di popolazioni. Il saltazionismo, quindi, fu sostituito da un processo graduale e la filosofia essenzialista da una nuova filosofia che non ammetteva più i tipi platonici, ma solo popolazioni variabili di individui tutti diversi fra loro. Anche se questo processo graduale è di gran lunga il più comune processo di speciazione, notiamo, di sfuggita, che casi di speciazione istantanea esistono, in particolare fra le piante, e sono prodotti da duplicazione cromosomica in specie ibride (allopoliploidia) e da altri processi cromosomici. Il verificarsi di questa speciazione istantanea, comunque, non sembra avere un gran significato evolutivo.
b) Speciazione.
Questo argomento è stato trattato in dettaglio in un altro articolo di questa Enciclopedia (v. specie e speciazione), pertanto in questa sede lo riassumeremo al massimo. Una condizione imprescindibile per capire la speciazione è aver capito la natura delle specie biologiche. Darwin sostituì al concetto di specie tipologica della scienza predarwiniana il concetto di specie di popolazioni. Secondo questo concetto le specie sono aggregati di popolazioni, isolati, dal punto di vista riproduttivo, da altri aggregati consimili. L'isolamento riproduttivo è attuato dai cosiddetti ‛meccanismi di isolamento' (come le barriere di sterilità o le incompatibilità comportamentali). Questi meccanismi di isolamento sono responsabili della discontinuità fra specie che condividono le stesse aree geografiche. Il problema della speciazione è come possa una tale discontinuità evolvere gradualmente.
Attualmente si concorda sul fatto che ciò si verifichi, nella maggioranza dei casi, per l'isolamento geografico delle popolazioni e la loro conseguente divergenza genetica. Questo processo si chiama ‛speciazione geografica' o ‛allopatrica' e si verifica in due forme: a) popolazioni che originariamente vivevano in contatto reciproco vengono separate da una barriera sorta in un secondo tempo, per esempio una catena montuosa, un braccio di mare o una discontinuità nella vegetazione. Mutazioni, processi stocastici e fattori di selezione saranno diversi nei due tronconi separati della specie originaria (essendo ogni scambio di geni fra di essi impedito dalla barriera geografica) e a tempo debito le due popolazioni figlie saranno sufficientemente diverse da comportarsi, l'una nei confronti dell'altra, come fossero due specie distinte; b) una popolazione fondatrice si stabilisce oltre i confini del territorio originario della specie. Questa nuova popolazione, fondata da una singola femmina fecondata, o da pochi individui, conterrà soltanto poche e spesso alquanto insolite combinazioni dei geni della popolazione madre, acquisterà una considerevole omozigosi a causa dell'inincrocio e sarà esposta a un nuovo insieme, probabilmente piuttosto severo, di pressioni di selezione, imposte dal mutato ambiente fisico e biotico. Una tale popolazione, quindi, viene modificata geneticamente in maniera alquanto drastica e può speciare rapidamente. Per di più una siffatta popolazione fondatrice, a causa della sua limitata base genetica e della drastica ristrutturazione genetica che subisce, risulta particolarmente adatta a dare origine a nuove linee evolutive (rivoluzione genetica). Questo tipo di speciazione all'interno di popolazioni fondatrici si chiama ‛speciazione peripatrica'.
Altri processi ipotizzati, come la ‛speciazione simpatrica' e ‛parapatrica', sono tuttora controversi e non chiaramente dimostrati (v. Barigozzi, 1982).
c) Discendenza comune.
Una delle più importanti teorie evolutive di Darwin era quella della discendenza comune. Darwin postulava che non solo tutte le specie di un genere discendessero da un'unica specie ancestrale, ma che, analogamente, tutti i generi di una famiglia, di un ordine, di una classe, di un phylum, in pratica tutti gli animali, discendessero in ultima analisi da un'unica specie ancestrale. Egli si spinse anche oltre e suggerì che tutte le forme di vita sulla Terra potessero aver avuto un'unica origine: ‟essendo stata infusa [la vita], in origine, in poche forme o in un'unica forma" (v. Darwin, 1859, p. 490). Questa non era altro che pura speculazione. Che animali e piante, per non citare i Batteri e altre forme di vita, potessero avere avuto un'origine comune sembrava, a tutta prima, assolutamente improbabile. Ma di lì a poco furono trovati dei protisti (organismi unicellulari), come certe alghe e certi flagellati, che possedevano tutte le caratteristiche postulate di un antenato comune ad animali e piante. Cosa ancora più importante, si scoprì che animali e piante hanno la stessa struttura cellulare, lo stesso sistema di trasmissione dei caratteri ereditari, la stessa composizione molecolare. Infine si scoprì persino che i Procarioti (Batteri e simili) hanno lo stesso codice genetico di animali e piante. Oggigiorno non sussiste più alcun dubbio che tutta la vita sulla Terra, qual è attualmente, abbia avuto un'unica origine.
La teoria della discendenza comune possiede uno straordinario potere esplicativo ed è quindi stata adottata da botanici e zoologi con grande entusiasmo. Essa spiegava le ragioni dell'esistenza della gerarchia linneana, spiegava le somiglianze anatomiche fra organismi appartenenti allo stesso tipo morfologico, spiegava la natura delle somiglianze fra le strutture molecolari di organismi più o meno strettamente imparentati. Per generazioni e generazioni, a partire dal 1859, zoologi e botanici sono stati impegnati a elaborare tutti i dettagli della discendenza. Anche se sussistono tuttora alcune incertezze, attualmente è del tutto chiaro che gli organismi possono essere raggruppati, fino a un certo punto principalmente per ragioni di convenienza, in cinque regni: 1, Monere (Procarioti); 2, Protisti (organismi unicellulari); 3, Funghi; 4, Metafiti (piante); 5, Metazoi (animali).
In base alla teoria della discendenza comune ci si aspetterebbe una completa continuità fra tutti i taxa di organismi. L'evoluzionista, a dire il vero, è convinto che una tale continuità ci sia stata, una volta, e che le piccole e grandi lacune che ora si riscontrano fra molti taxa siano dovute al fatto che diverse specie si sono estinte. Che una tale estinzione abbia effettivamente avuto luogo è ben documentato nei più recenti strati geologici, che contengono probanti reperti fossili. Per quel che riguarda i Vertebrati, gli anelli mancanti sono rappresentati da Archaeopteryx fra Rettili e Uccelli, dai rettili tipo mammiferi fra Rettili e Mammiferi, da Jchthyostega fra Pesci e Anfibi, ecc. Non ogni filogenesi è stata divergente. Nelle piante l'ibridazione può talvolta portare a nuove specie e attualmente si dispone di molte prove che gli Eucarioti hanno avuto origine da una simbiosi fra linee di Procarioti preesistenti. Comunque quasi tutta l'evoluzione nell'ambito degli Eucarioti è stata divergente.
d) L'origine della vita.
Nel XVIII secolo e nei primi anni del XIX i più credevano nella generazione spontanea, cioè nel fatto che la vita traesse origine dalla materia inanimata. Questa credenza fu distrutta ai tempi di Darwin attraverso le ricerche di L. Pasteur e di altri microbiologi. Eppure, per Darwin e per i suoi seguaci, non v'era dubbio che, all'inizio, la vita sulla Terra dovesse essere sorta spontaneamente. Oggi si sa che la vita non potrebbe avere origine in un'atmosfera ricca di ossigeno qual è l'attuale atmosfera terrestre. Ma disponiamo di sicuri indizi che da tre miliardi e mezzo a quattro miliardi di anni fa, quando ebbe origine la vita, la Terra avesse un'atmosfera riducente, ricca di metano, di ammoniaca e di gas analoghi, ma totalmente priva di ossigeno.
Anche se alcuni passaggi sono ancora incerti, siamo ora in grado di ricostruire un ragionevole ‛scenario' dell'origine della vita a partire da precursori presenti in natura, come gli amminoacidi e gli acidi nucleici (v. vita, origine della; v. Küppers, 1983). Un fatto particolarmente interessante è che dalle ricostruzioni più recenti risulta che già nello stadio prebiotico abbia svolto un ruolo importante una forma di selezione naturale.
e) Mutamento evolutivo e adattamento.
Lo studio del mondo vivente indusse molti naturalisti religiosi, nei secoli XVII e XVIII, a esprimere la loro ammirazione per la meravigliosa struttura del mondo. Tutti gli organismi sembrano perfettamente adattati al posto che occupano in natura e agli altri organismi con cui convivono. Si definì ‛teologia naturale' lo studio della natura teso a conoscere meglio la bontà e la saggezza di Dio profuse nella concezione del mondo. Darwin e altri naturalisti, da Lamarck in poi, studiarono le stesse meraviglie dell'adattamento, ma tentarono di spiegarle in termini naturali. Questo tentativo affronta il secondo grande problema dell'evoluzione, quello dell'origine dell'adattamento e dell'apparente reciproco adattamento di tutte le parti degli organismi.
La teoria della selezione naturale di Darwin aveva come obiettivo una spiegazione in termini naturali dell'adattamento e dell'armonia complessiva del mondo vivente. Il ragionamento di Darwin era il seguente: esiste una grande sovrapproduzione di individui a ogni generazione, ma, dal momento che le risorse naturali sono limitate, soltanto una piccolissima percentuale dei membri di ogni generazione può sopravvivere. In secondo luogo, tutti i nati differiscono l'uno dall'altro per quel che riguarda il patrimonio genetico e quindi, almeno in linea di principio, differiscono per quel che riguarda il rispettivo adattamento all'ambiente comune e l'abilità di competere con i propri simili. In terzo luogo, le cause delle differenze nelle capacità di adattamento sono in parte ereditarie. Ne consegue, per semplice logica, che gli individui dotati delle maggiori capacità di adattamento hanno maggiori possibilità di sopravvivere e di riprodursi. In virtù di questa sopravvivenza non casuale vi sarà un mutamento continuo nel mondo vivente, in altre parole vi sarà evoluzione.
Benché apparentemente così logica, la teoria della selezione naturale di Darwin fu, in un primo tempo, quasi completamente respinta. Essa rappresentava una rottura troppo drastica con il modo di pensare allora vigente. Ci vollero quasi ottant'anni prima che la teoria della selezione naturale fosse adottata da tutti gli evoluzionisti, e tuttora si riscontra una grande resistenza contro questa teoria, al di fuori del campo della biologia evolutiva. Quando si leggono gli argomenti sollevati contro la validità della teoria della selezione naturale, ci si rende subito conto che, in massima parte, sono basati su gravi incomprensioni. Per questo motivo è importante spiegare in dettaglio la struttura della teoria della selezione naturale.
f) La selezione naturale é un processo a due stadi.
Il primo stadio della selezione naturale consiste nei processi che generano la moltitudine di individui, i quali nel secondo stadio sono esposti alla selezione naturale. Dato che ognuno di questi individui è unico, dal punto di vista genetico, si può anche dire che il primo passo della selezione naturale consiste nella produzione di variazione genetica. Che questa variazione si producesse di continuo era del tutto evidente per Darwin, e l'intera sua teoria poggiava sul riconoscimento di questo fatto. Tuttavia i meccanismi che producevano questa variazione costituivano per Darwin un mistero assoluto. Il problema è stato alla fine risolto, dopo il 1900, dalla genetica (v. evoluzione: La moderna teoria dell'evoluzione).
Nel corso della meiosi, un processo cellulare che costituisce una fase della gametogenesi, i cromosomi paterni e materni vanno incontro a ricombinazione genica e nella successiva divisione riduzionale solo un cromosoma, per ogni coppia di cromosomi omologhi dei genitori, entra a far parte della cellula che diventa un gamete. Si ha un'altissima sovrapproduzione di gameti, in particolare di gameti maschili, e l'accoppiamento di individui maschi e femmine, nonché la selezione di quei particolari gameti che si uniscono nella fecondazione, sono in larghissima misura fatti casuali. In altre parole, tutti i processi che precedono la fecondazione, dal crossing-over dei cromosomi paterni e materni alla loro separazione durante la divisione riduzionale, dalla scelta del partner all'incontro fra i due gameti che costituiranno lo zigote, sono eventi interamente o, almeno, prevalentemente casuali. Le possibilità di produzione di variazione durante questo primo stadio del processo di selezione naturale sono illimitate.
Con la produzione di uno zigote, un uovo fecondato, comincia il secondo stadio del processo di selezione naturale. Da questo momento in poi la ‛buona qualità' del nuovo individuo è costantemente messa alla prova, dallo stadio di larva (o di embrione) fino all'età adulta e al periodo riproduttivo. Solo gli individui più efficienti nel far fronte alle avversità dell'ambiente e agli scontri con gli altri membri della stessa specie sopravviveranno fino all'età della riproduzione e si riprodurranno con successo. L'esperienza ci ha insegnato che in questo secondo stadio del processo di selezione naturale certi individui dotati di particolari patrimoni genetici sono chiaramente superiori agli altri. È questa osservazione che ha indotto a coniare l'espressione, un po' ambigua, ‛sopravvivenza del più idoneo'. La differenza fondamentale fra il primo e il secondo stadio della selezione naturale dovrebbe ora essere chiara: nel primo stadio, quello della produzione di variazione genetica, ogni evento è casuale; nel secondo stadio, quello della sopravvivenza e della riproduzione differenziali, il caso svolge un ruolo molto minore e il successo dell'individuo è in larga misura determinato dalle sue caratteristiche fondate geneticamente. Dire che la selezione naturale è un processo casuale rivela una totale incomprensione del fenomeno.
La produzione di variazione, il primo passo del processo di selezione naturale, ha costituito uno dei più importanti problemi per gli evoluzionisti, fin dalla nascita della genetica. Prima dell'avvento della sintesi evolutiva si pensava che la principale fonte di variazione fosse la pressione di mutazione. A tempo debito, comunque, si capì che la principale fonte del materiale che viene offerto alla selezione naturale è la ricombinazione dei genotipi dei due genitori; pertanto la formula, spesso usata, ‟l'evoluzione è il risultato di mutazione e selezione" e erronea. Benché, naturalmente, la mutazione sia la causa ultima di tutte le differenze genetiche, il materiale su cui effettivamente si esercita la selezione è costituito dagli individui prodotti attraverso la ricombinazione: non esistono, infatti, due fratelli o due sorelle (a parte i gemelli omozigoti) geneticamente identici.
Questa variazione è ‛casuale' in due sensi: a) non è prevedibile; b) non costituisce la risposta a una qualche necessità dell'organismo in cui si verifica. Ma ‛casuale' non vuol dire che ogni concepibile variazione sia possibile; al contrario il grado di variazione è fortemente vincolato dal genotipo (v. oltre). Questi vincoli sono imposti dalla sequenza di coppie di basi del genotipo e, dopo la sua traduzione, dalla capacità della nuova proteina di collaborare con il resto dell'apparato di sviluppo.
In anni recenti, grazie alla biologia molecolare, è stato scoperto un altro tipo di variazione, che si potrebbe chiamare ‛variazione distorta'. L'esempio più vistoso di questa variazione è la variazione prodotta da elementi trasponibili. Questi elementi hanno la capacità di replicarsi indipendentemente dal ciclo meiotico e si inseriscono in altri cromosomi; alcuni di essi esistono in molte centinaia di migliaia di copie. Questi frammenti di DNA altamente replicati pongono due problemi. Primo: perché la selezione naturale non è in grado di evitare una tale replicazione, apparentemente inutile, che consuma le risorse metaboliche di un organismo? Certo questa replicazione sarebbe soppressa dal meccanismo selettivo non appena inficiasse l'idoneità del fenotipo, ma, a giudicare dalla sua frequenza, si deve concludere che, per lo più, deve risultare neutrale.
Il secondo problema è come mai la disponibilità di un tale sovrappiù di DNA non influenzi il tasso di mutazione in maggior misura. Come ora ben si sa, molte specie, forse le più diffuse, godono di grande longevità, misurabile in 5, 10 o persino più di 20 milioni d'anni. Perché tutto questo sovrappiù di DNA non accelera l'evoluzione? Ciò conferma forse il fatto che questo DNA (non codificante?) è del tutto neutrale?
Talvolta si definisce la produzione vincolata e distorta di variazione come una forma di selezione. Questa designazione è ingannevole: la produzione vincolata e distorta di variazione può in effetti influenzare la direzione del mutamento evolutivo, ma certamente non è una forma di selezione.
g) Il bersaglio della selezione.
Darwin non aveva dubbi sul fatto che il bersaglio della selezione fosse l'individuo: è l'individuo che sopravvive o meno, che si riproduce con successo o no. Tuttavia, nel periodo riduzionista, durante gli anni venti, i genetisti matematici delle popolazioni scelsero, invece, il gene come bersaglio della selezione. Certo può darsi che un singolo gene sia responsabile della superiorità o dell'inferiorità di un individuo, ma ciò non toglie che sia l'individuo nel suo insieme, o, più correttamente, il suo fenotipo, il bersaglio effettivo della selezione. Una volta capito chiaramente questo fatto, alcune grosse difficoltà, inerenti al punto di vista del ‛gene come bersaglio della selezione', scompaiono. Per esempio, la ricerca nel campo della genetica molecolare ha dimostrato che molte mutazioni (sostituzioni di alleli) sono ‛neutrali', non hanno, a quanto risulta, valore selettivo. In realtà la maggior parte dei genotipi contiene addirittura una certa percentuale di geni moderatamente nocivi. La presenza di questi geni nelle popolazioni non dimostra l'invalidità del principio di selezione naturale di Darwin, perché essi sono semplicemente trasportati come ‛autostoppisti' da genotipi per il resto validi. Lo stesso discorso vale a livello fenotipico: spesso certi aspetti del fenotipo sono conservati nel corso dell'evoluzione anche se non contribuiscono alle capacità di adattamento degli individui in cui compaiono; sono tollerati fintantoché non interferiscono con l'adattamento complessivo degli individui in cui si manifestano.
h) Selezione di sopravvivenza.
Evidentemente nessun individuo può contribuire al pool genetico della generazione successiva se non sopravvive fino all'età della riproduzione; il tasso di mortalità fra il momento della fecondazione dello zigote e l'età riproduttiva è molto alto. Dato che ogni individuo, nelle specie che si riproducono sessualmente, è diverso da tutti gli altri membri della popolazione cui appartiene, in media differirà dagli altri per numerose proprietà: per la sua capacità di resistere agli agenti patogeni e di sfuggire ai nemici, per la sua abilità nel procurarsi il cibo e per numerose altre capacità di adattamento che ne faciliteranno la sopravvivenza. Gran parte della mortalità fra il momento della fecondazione e l'età adulta è dovuta all'eliminazione di individui (zigoti) che difettano di, o posseggono in misura non adeguata, l'una o l'altra capacità adattativa. Ciò corrisponde all'eliminazione preevolutiva di ‛degenerazioni del tipo'. Non tutta la selezione è di questo tipo negativo: la selezione naturale non solo colpisce genotipi inferiori, ma favorisce anche la sopravvivenza e la riproduzione di genotipi superiori. Dato che questi individui favoriti sono il risultato di una ricombinazione genica che si verifica durante la meiosi, la selezione naturale viene spesso considerata, e in questo caso piuttosto giustamente, un processo creativo: è questo processo creativo che dà come risultato l'adattabilità degli individui.
i) Adattamento.
A prima vista niente parrebbe più semplice del concetto di adattamento: si potrebbe dire che un organismo è ben adattato se possiede i tratti necessari a sopravvivere con successo. Ma come stabilire un fatto del genere? Senza dubbio i Pesci sono ben adattati a vivere nell'acqua; eppure sappiamo, grazie ai reperti fossili, che diecimila o più specie di Pesci si sono estinte. Altre specie sono sopravvissute e hanno prosperato perché ‛meglio adattate': ma cosa significa questo? I tratti adattativi di un organismo sono molti e diversi: non comprendono solo la capacità di tollerare l'ambiente fisico (il clima), ma anche le capacità di difesa contro le malattie (gli agenti patogeni) e contro i nemici, e la capacità di competere vittoriosamente con altri individui per procacciarsi varie risorse dell'ambiente, come il cibo e il territorio. È evidente, quindi, che un organismo deve essere adattato in molti modi diversi. Alcune necessità adattative sono reciprocamente incompatibili e la selezione realizza un compromesso fra loro; una delle ragioni per cui nessun organismo è perfetto è che un compromesso non può mai raggiungere la perfezione; altre ragioni che determinano questa mancanza di perfezione sono i molteplici limiti di cui parleremo più avanti.
Si è tentato in molti modi di misurare l'adattamento. Spesso si afferma che il grado di adattamento si riflette nel contributo relativo (in termini di prole) alla generazione successiva; in molti casi questo criterio fornisce una misura adeguata, ma consideriamo il caso di due gemelli identici che camminano in una foresta durante un temporale: uno dei due viene ucciso da un fulmine, mentre l'altro sopravvive e genera otto figli. Si può affermare che il gemello con otto figli fosse meglio adattato? Certamente no, in senso biologico. Considerazioni come questa hanno portato a concepire l'adattamento come ‛tendenza' a sopravvivere e a riprodursi con successo. È la ‛speranza' (in senso statistico) in questo successo, tolti tutti gli incidenti e le evenienze improbabili, che misura il grado di adattamento.
Il concetto di adattamento implica che l'adattamento non è un fatto accidentale, ma il risultato di un processo selettivo avvenuto nel corso delle generazioni precedenti, ovvero il risultato di una selezione che ha ulteriormente perfezionato le primitive versioni dei tratti adattativi presenti nell'organismo attualmente vivente: l'adattamento è sempre il prodotto finale di una storia passata.
Dal momento che il bersaglio della selezione è l'individuo nel suo complesso, questi è considerato ben adattato se riesce ad affermarsi nel corso del processo di selezione. Singole componenti del fenotipo non sono sottoposte alla selezione separatamente, ma solo in quanto parti dell'intero fenotipo; per questo motivo non è necessario che singoli aspetti del fenotipo siano perfetti, fintantoché il fenotipo nel suo insieme risulta ben adattato.
L'aver sottolineato il fatto che l'adattamento è il prodotto di una serie di eventi selettivi passati è importante quando si passa a considerare le modifiche che intervengono nel corso del processo di adattamento. Gli eventi evolutivi più importanti sono gli spostamenti di certi organismi in nuove nicchie o in nuove zone adattative. Quando gli antenati dei Tetrapodi, i primi anfibi, emersero dall'acqua, non erano affatto perfettamente adattati alla vita sulla terraferma; comunque possedevano certe strutture, come i polmoni, uno scheletro interno, estremità che consentivano loro di camminare, ecc., che costituivano altrettanti preadattamenti alla vita sulla terraferma. Ciò dimostra che alcune strutture o altre caratteristiche di un organismo, oltre a costituire adattamenti al particolare ambiente in cui l'organismo vive al momento, possono anche assumere il ruolo di preadattamenti quando l'organismo in questione si sposta in una zona adattativa diversa.
Darwin, sulle prime, pensò che la selezione naturale portasse a un adattamento perfetto; ma ben presto si rese conto di quanto fossero imperfetti quasi tutti gli organismi. Questo fatto non sorprende, se si considera la natura del processo di selezione attraverso cui si acquistano i caratteri adattativi. Non si capirà mai il concetto darwiniano di evoluzione se non ci si rende conto che l'evoluzione darwiniana non è un processo teleologico, diretto a un fine; nè è un processo deterministico, non obbedisce ad alcuna ‛necessità': l'evoluzione è sempre opportunistica e il suo successo è solo probabilistico.
Malgrado la casualità del processo di selezione naturale, gli organismi che vivono in un ambiente più o meno stabile sono, in genere, ragionevolmente ben adattati. Comunque ogni organismo è messo alla prova a ogni generazione e soltanto pochi membri di ogni generazione riescono a riprodursi generando i membri della generazione successiva; e dato che a ogni generazione la costellazione delle forze selettive, nonché la natura dei singoli genotipi mutano, sarà diversa, di conseguenza, la risposta a una nuova pressione selettiva: questo è il motivo per cui la selezione non è un processo deterministico nè finalistico.
Quando l'evoluzionista studia un organismo, cerca di ricostruire le forze selettive che hanno portato alla formazione delle sue diverse caratteristiche. Perché la specie a è verde, mentre la specie b è marrone? Perché la specie a è liscia e la specie b ha le spine? Perché in questa famiglia di Mammiferi le femmine hanno le corna, mentre in un'altra famiglia le hanno soltanto i maschi? Queste differenze hanno un significato adattativo? Il cosiddetto ‛programma adattamentista' consiste proprio nel porsi questo tipo di domande. Questa strategia di ricerca ha conseguito straordinari successi e ha permesso di capire molte caratteristiche che prima non si riusciva a spiegare, per esempio la funzione dell'organo della linea laterale dei Pesci: il chiedersi il perché ha costituito un ottimo strumento euristico. Il fatto di essersi chiesto perché le vene abbiano delle valvole aiutò Harvey a scoprire la circolazione del sangue. Comunque il programma adattamentista nasconde dei rischi: dal momento che il bersaglio della selezione è l'organismo nel suo insieme, sarebbe un errore insistere sul fatto che ogni singola caratteristica di un organismo debba possedere un proprio valore adattativo. Il programma adattamentista va, quindi, applicato con cautela.
3. Limiti all'efficacia della selezione naturale.
Anche se ‟la selezione naturale esamina e controlla di giorno in giorno e di ora in ora, in ogni parte del mondo, qualsiasi variazione, anche la più piccola", come dichiarò Darwin (v., 1859, p. 84), è tuttavia evidente che esistono limiti severi all'efficacia della selezione. Niente dimostra questo stato di cose in maniera più convincente del fatto che il 99,9% più di tutte le linee evolutive si è estinto. Dobbiamo quindi chiederci perché la selezione naturale si dimostri così spesso incapace di produrre la perfezione. Studi recenti hanno rivelato che le ragioni di questa limitazione sono molte. Discutere questi limiti nei particolari contribuisce molto a capire l'evoluzione.
1. Il potenziale del genotipo. - L'effettiva organizzazione genetica di un animale o di una pianta impone severi limiti alla sua ulteriore evoluzione. Come ha detto Weismann, nessun uccello potrà mai evolversi in un mammifero e nessuno scarafaggio in una farfalla. Gli Anfibi non sono stati capaci di generare una discendenza in grado di vivere nell'acqua salata. Noi ci meravigliamo che i Mammiferi siano stati capaci di sviluppare organi per il volo (pipistrelli) e per la vita nell'acqua (cetacei e foche), ma esistono molte nicchie ecologiche che i Mammiferi non sono stati in grado di occupare. Per esempio, le loro dimensioni non possono scendere al di sotto di un certo limite, e la selezione, per quanto massiccia, non è riuscita a produrre mammiferi più piccoli del toporagno nano.
2. Assenza di variazione genetica appropriata. - Una data specie o una data popolazione può tollerare solo una dose molto limitata di variazione. Nel caso di un qualche drastico mutamento dell'ambiente, che si tratti di un deterioramento delle condizioni climatiche o della comparsa di un nuovo nemico o concorrente, il tipo di geni necessari per fornire una risposta appropriata a questa nuova pressione selettiva può non essere presente nel pool genetico della popolazione; l'alta frequenza di estinzione documenta l'importanza di questo fattore.
3. Processi stocastici. - Gran parte della sopravvivenza e della riproduzione differenziali in una popolazione non dipende dalla selezione, bensì dal caso. Il caso opera a ogni livello del processo riproduttivo, a cominciare dal crossing-over fra i cromosomi paterni e materni, durante la meiosi, fino alla sopravvivenza degli zigoti di nuova formazione. Non vi è alcun dubbio che combinazioni di geni potenzialmente favorevoli siano spesso distrutte da forze ambientali che agiscono indiscriminatamente, come alluvioni, terremoti, eruzioni vulcaniche, senza che la selezione naturale abbia l'opportunità di favorire questi genotipi.
4. Percorsi multipli. - In genere a una sfida lanciata dall'ambiente è possibile fornire diverse risposte alternative: è l'effettiva struttura dell'organismo che spesso determina la soluzione adottata. Allorché per gli antenati dei Vertebrati e per quelli degli Artropodi rappresentò un vantaggio evolutivo lo sviluppo di uno scheletro, gli antenati degli Artropodi possedevano i prerequisiti per sviluppare uno scheletro esterno, quelli dei Vertebrati uno scheletro interno. Tutta l'evoluzione di questi due grandi gruppi di organismi è stata fin d'allora influenzata da queste due diverse scelte operate dai rispettivi antenati. La soluzione adottata dai Vertebrati permise lo sviluppo di enormi creature, come i dinosauri, gli elefanti e le balene, mentre il più grande tipo di artropodo che sia mai esistito è un grosso granchio. La necessità di una muta regolare dello scheletro esterno impone una potente pressione selettiva contro l'aumento di dimensioni. Le specie di nuotatori pelagici, discese da antenati sessili o bentonici o nuotatori attivi, appartenenti a diversi taxa animali, si sono adattate alla forma di vita pelagica attraverso percorsi evolutivi completamente diversi. Ogni evoluzione rappresenta un diverso compromesso fra i requisiti necessari per invadere la nuova zona adattativa e la struttura preesistente. Invariabilmente, durante l'evoluzione, l'adozione di una soluzione particolare di fronte a una nuova opportunità ambientale restringe notevolmente le possibilità di evoluzione futura.
5. Capacità di modificazione non genetica. - Più flessibile è il fenotipo (grazie alla flessibilità del processo di crescita), più ciò riduce la forza di una pressione selettiva avversa. Le piante, e in particolare i microrganismi, hanno una capacità di modificazione fenotipica di gran lunga maggiore rispetto agli animali superiori. La capacità di modificazione non genetica è comunque presente persino nell'Uomo, come dimostrano, per esempio, i cambiamenti fisiologici che avvengono in una persona che si sposti da una zona di pianura a una zona di alta montagna: col passare dei giorni e delle settimane il suo organismo può adattarsi ragionevolmente bene alla pressione atmosferica più bassa. La selezione naturale, ovviamente, interviene anche in questo fenomeno, dal momento che la capacità di adattamento non genetico è sotto stretto controllo genetico. Succede anche che, se una popolazione si sposta in un nuovo ambiente con caratteristiche specifiche, nel corso delle generazioni successive vengano selezionati geni che rinforzano e alla fine rimpiazzano in larga misura la capacità di adattamento non genetico.
6. Interazione durante lo sviluppo. - Alcuni studiosi di morfologia, a partire da Étienne Geoffroy Saint-Hilaire, si sono accorti che esiste competizione fra organi e strutture. Geoffroy espresse questo fatto nella ‟legge del bilanciamento" (loi de balancement). Le diverse componenti del morfotipo non sono indipendenti l'una dall'altra e nessuna di esse risponde alla selezione senza interagire con le altre. L'intero apparato dello sviluppo costituisce un unico sistema interagente; gli organismi rappresentano compromessi fra richieste contrastanti. Fino a che punto una particolare struttura o un organo possa rispondere alle forze della selezione dipende in larga misura dalla resistenza opposta da altre strutture e da altre componenti del genotipo. Wilhelm Roux, circa cento anni fa, definì le interazioni competitive durante lo sviluppo ‟lotta delle parti" negli organismi.
La morfologia di ogni organismo mostra fino a che punto essa sia il risultato di un compromesso. Ogni cambiamento di zona adattativa lascia un residuo di caratteristiche morfologiche che costituiscono, in effetti, un impedimento. Basta ricordare i molti punti deboli dell'uomo, che sono i resti del suo passato di quadrupede e di animale più vegetariano: per esempio i seni facciali, la struttura della parte caudale della colonna vertebrale e l'appendice cecale. Molti tratti caratteristici dell'ontogenesi, che vengono generalmente citati come prove della ricapitolazione, rientrano nella stessa categoria. Per un embrione di mammifero o di uccello il fatto di dover passare attraverso lo stadio di formazione dell'arco branchiale, durante l'ontogenesi, sembrerebbe un modo molto poco efficiente per costruire la regione del collo. Eppure il complesso delle interazioni nel corso del processo di sviluppo è così ben integrato che la selezione naturale finora non è stata capace di trovare una scorciatoia per un percorso più diretto.
7. La struttura del genotipo. - La classica metafora usata per descrivere il genotipo è quella della collana di perle, nel senso che i geni sarebbero allineati uno appresso all'altro come le perle di una collana. Secondo questo classico punto di vista, ogni gene sarebbe più o meno indipendente dagli altri e tutti i geni sarebbero di natura più o meno simile. Questa concezione, tanto in voga cinquant'anni fa, è stata quasi del tutto abbandonata. Certo tutti i geni sono fatti di DNA e l'informazione che contengono è codificata nella sequenza di paia di basi; tuttavia le più recenti ricerche nel campo della biologia molecolare suggeriscono l'ipotesi che esistano diverse classi funzionali di geni, alcune col compito di produrre materiale, altre di regolarne la produzione (v. gene, suppl.). Inoltre esiste un buon numero di prove indirette del fatto che molti geni sono organizzati in sistemi funzionali, che per molti versi agiscono all'unisono. Comunque questo è forse il settore più controverso della biologia evolutiva e, per ora, la cosa migliore da fare è richiamare l'attenzione sul fatto che la vecchia concezione del genotipo come ‛collana di perle' non è più considerata valida, ma sussiste ancora grande incertezza a proposito dell'effettiva organizzazione del genotipo. Il fatto che esistano geni monocodificanti (single-coding), DNA a media ripetitività, DNA ad alta ripetitività, transposoni, esoni e introni, oltre a molti altri tipi di DNA, suggerisce l'esistenza di funzioni differenziali, ma quali siano e come cooperino l'una con l'altra resta ancora da scoprire. Da quel che capiremo sulla struttura del genotipo impareremo, sul modo di operare dell'evoluzione, più che da qualsiasi altro tipo di ricerca.
4. Estinzione.
Alla moltiplicazione delle specie (speciazione) si contrappone il fenomeno dell'estinzione. L'estinzione del dodo e del colombo migratore sono esempi di estinzioni avvenute in tempi recenti, ma i reperti fossili contenenti i resti di milioni di organismi che non esistono più indicano che l'estinzione è un fenomeno che, a quanto pare, risale all'origine della vita sulla Terra. I due problemi più importanti, riguardo all'estinzione, sono: primo, ‛quali sono le cause dell'estinzione?', e, secondo, ‛l'estinzione è un processo continuo o si verifica in brevi periodi caratterizzati da eventi catastrofici?'. Cuvier, Louis Agassiz e molti geologi del primo Ottocento credevano nell'estinzione ‛catastrofica', ma dopo il 1859 l'opinione prevalente era che il tasso di estinzione fosse circa lo stesso in tutte le epoche. Attualmente si propende per una concezione che rappresenta una sintesi delle due opposte opinioni: certo l'estinzione si verifica in tutte le epoche, a causa della competizione fra specie diverse e dei lenti cambiamenti nelle faune e nelle flore, cui alcune specie sono incapaci di far fronte; ma ora possediamo anche le prove che ci sono stati dei periodi di estinzione catastrofici, di cui il meglio documentato è il periodo di transizione dal Cretaceo al Terziario. Possediamo validi indizi del fatto che la Terra, in quell'epoca, fu colpita da un asteroide, che probabilmente produsse una nuvola di polvere così densa da dar luogo a un temporaneo oscuramento dell'atmosfera e a un letale raffreddamento della superficie terrestre. Molti gruppi di organismi, come, per esempio, i dinosauri, soccombettero a questo evento, mentre altri, come i Mammiferi, sopravvissero e, per un certo periodo, furono sottoposti a una notevole quantità di radiazioni nel nuovo biotopo. Non si è ancora capito perché alcuni gruppi si estinguano e altri sopravvivano. Per quel che riguarda altri periodi di estinzione, in ere geologiche remote, il fenomeno va attribuito a un raffreddamento della Terra o a qualche altro drastico cambiamento di clima; ma attualmente il fattore di estinzione di gran lunga più importante è l'uomo, particolarmente in quanto responsabile del rapido disboscamento dei tropici.
5. Il ruolo del comportamento.
Per Lamarck il comportamento era un'importante causa di mutamento evolutivo. Egli riteneva che i cambiamenti prodotti negli organismi da un qualsiasi tipo di attività fossero trasmessi alle generazioni successive tramite una forma di ereditarietà dei caratteri acquisiti. Anche se la teoria di Lamarck oggi viene respinta, noi crediamo ancora che il comportamento sia importante nell'ambito dell'evoluzione, ma in un senso completamente diverso. Cambiamenti nel comportamento, per esempio l'adozione di nuove abitudini alimentari o la diffusione di una popolazione su un territorio più vasto, possono far insorgere nuove pressioni selettive, che, a loro volta, possono portare a mutamenti evolutivi. Vi è motivo di credere che cambiamenti di comportamento siano stati fra i fattori che hanno determinato gran parte delle innovazioni evolutive. Lo stesso comportamento, d'altronde, possiede una grossa componente genetica ed è soggetto al mutamento evolutivo come i caratteri strutturali. Lo studio dell'evoluzione e del significato selettivo di componenti del comportamento è diventato, in anni recenti, uno dei più importanti settori della biologia del comportamento.
6. Selezione sessuale.
La selezione, come l'abbiamo descritta fin qui, potrebbe portare a una maggiore probabilità di sopravvivenza, determinata dal migliore adattamento all'ambiente fisico e biotico. Questo miglior adattamento comprenderebbe tutti gli aspetti di una maggiore efficienza fisiologica, la capacità di affrontare con successo predatori e rivali, una particolare flessibilità comportamentale in situazioni di emergenza, e qualità simili. Tuttavia non sempre la selezione porta a un miglior adattamento: può darsi che un individuo fornisca un maggior contributo di geni alla generazione successiva semplicemente perché ha più successo nel riprodursi. Darwin chiamò questo tipo di selezione ‛selezione sessuale'. Un qualsiasi carattere di un maschio che porti a un maggior successo riproduttivo, per esempio la capacità di fecondare più femmine, sarà automaticamente favorito dalla selezione, a meno che non sia contrastato da un qualche attributo avverso. Certi caratteri sessuali secondari maschili, come, per esempio, le penne degli uccelli del paradiso, sono tradizionalmente citati come esempi di fattori di selezione sessuale, ma anche molti aspetti del comportamento riproduttivo, in particolar modo dei maschi, rientrano in questa categoria. Al processo di selezione sessuale contribuisce la cosiddetta ‛scelta da parte della femmina', cioè la tendenza delle femmine a non accettare indiscriminatamente un qualsiasi maschio come partner sessuale, ma a mostrare ben definite preferenze.
Nel caso del comportamento sessuale è particolarmente facile constatare che la selezione è ‛egoista', il che rappresenta una necessità quasi inevitabile, considerato che il bersaglio della selezione è l'individuo. Perciò fu ben presto sollevata la questione se un comportamento altruistico potesse essere spiegato alla luce della selezione naturale e, in caso contrario, se non costituisse un controesempio nei confronti del darwinismo. Perché ci si chiedeva un individuo dovrebbe fare dei sacrifici o correre dei rischi allo scopo di procurare un vantaggio riproduttivo a un'altra persona? In effetti ciò sembrava, a prima vista, alquanto implausibile; ma Haldane, Hamilton, Williams, Trivers e altri hanno mostrato che il fatto di aiutare parenti stretti è in effetti favorito dalla selezione naturale, in quanto individui strettamente imparentati hanno, in parte, lo stesso genotipo: se un individuo contribuisce alla sopravvivenza dei propri parenti stretti, in effetti egli favorisce la sopravvivenza del proprio genotipo (selezione di parentela). Che un tale altruismo fra parenti esista risulta evidente dal comportamento di molti organismi nei confronti dei coniugi e dei figli; spesso però anche nei confronti di parenti più lontani viene adottato un comportamento altruista, per esempio emettendo segnali d'allarme.
Ancor più interessanti sono i casi di altruismo reciproco, come quelli osservati, per esempio, fra i babbuini: un individuo ne aiuta un altro, il quale, a sua volta, ricambia il favore in una diversa occasione. Il progresso fondamentale nell'etica umana dev'essersi verificato quando, per la prima volta, un individuo ha dimostrato dell'altruismo nei confronti di persone non legate a lui da vincoli di parentela. L'altruismo si riscontra in particolar modo fra gli animali sociali. Lo studio delle cure parentali e di altre forme di altruismo, alla luce della teoria della selezione naturale, costituisce attualmente un fertile campo di ricerca.
D'altra parte l'evoluzionista moderno non crede più che la selezione agisca ‛per il bene della specie': agisce solo per il bene degli individui, e soltanto attraverso di loro, molto indirettamente, per il bene della specie. Per un certo periodo fra gli evoluzionisti ha goduto di un certo credito la teoria della ‛selezione di gruppo', una teoria in base alla quale beneficiano della selezione era considerato il gruppo nel suo insieme. Ma un'analisi più approfondita del problema ha posto in tali difficoltà questa teoria, che la si è dovuta abbandonare quasi del tutto. Nei casi in cui, apparentemente, la teoria della selezione di gruppo sembrava confermata, si è quasi sempre potuto constatare che il gruppo consisteva di individui imparentati fra loro: si trattava quindi di casi di selezione di parentela (idoneità inclusiva).
Alcuni autori moderni hanno anche individuato una forma di ‛selezione di specie'. Fin dai tempi di Lyell e di Darwin si sa che quando una specie apparentemente meglio adattata compare in una determinata zona, altre specie possono estinguersi. Ogni volta che una linea filetica, per esempio quella che ha portato dagli insettivori ai pipistrelli, o dai Rettili agli Uccelli, ha occupato una nuova zona adattativa, le specie che sono riuscite meglio a realizzare rapidamente il mutamento adattativo hanno indotto l'estinzione degli anelli intermedi meno efficacemente adattati. In questo senso, in effetti, una selezione di specie esiste; ma ciò non è in contrasto con la selezione individuale. Gli individui delle due specie in competizione occupano, per così dire, un'unica nicchia ecologica, nella quale alcuni individui hanno più successo di altri. Anche se la maggior parte degli individui che sopravvivono appartiene a una specie e la maggior parte di quelli che soccombono all'altra specie, il risultato è sempre dovuto alla selezione individuale.
Si è anche sostenuto che esista una forma di selezione di specie quando le specie appartenenti a una linea filetica speciano più rapidamente di quelle di un'altra linea. Di fronte all'estinzione - si è detto - la linea più ricca di specie avrebbe una maggiore probabilità di sopravvivere dell'altra linea. In media questo puo, effettivamente, essere vero, anche se per indicare questo processo potrebbe essere più adatta l'espressione ‛selezione di speciazione'; inoltre, tutte le caratteristiche che favoriscono una rapida speciazione (capacità di diffusione e di colonizzazione, tendenza all'acquisizione di meccanismi di isolamento, ecc.) sono basate su proprietà genetiche di individui. Pertanto anche questo tipo di selezione di specie si fonda sulla selezione individuale.
7. Macroevoluzione.
Conviene dividere i fenomeni evolutivi in due categorie: la microevoluzione e la macroevoluzione. La microevoluzione concerne la variazione nelle popolazioni e la speciazione: in breve tutti i fenomeni e i processi che avvengono al livello e al di sotto del livello della specie. La macroevoluzione riguarda i processi che si verificano al di sopra del livello della specie, in particolare l'origine di nuovi taxa superiori, l'invasione di nuove zone adattative e, in correlazione con quest'ultimo fenomeno, l'acquisizione, che spesso si determina, di nuove caratteristiche evolutive, come le ali degli Uccelli, l'adattamento alla vita terrestre dei Tetrapodi e il sangue caldo negli Uccelli e nei Mammiferi.
La relazione fra microevoluzione e macroevoluzione è stata oggetto, recentemente, di notevoli discussioni e controversie. Le incertezze al riguardo possono essere in buona misura chiarite tenendo presente il fatto che tutti gli eventi macroevolutivi hanno luogo all'interno di popolazioni e nel genotipo di singoli individui, e quindi sono sempre, contemporaneamente, processi microevolutivi. Nello stesso tempo esiste, in effetti, una netta differenza fra macro- e microevoluzione a livello fenotipico; pertanto i fenomeni macroevolutivi richiedono la formulazione di ipotesi e modelli specifici. Gli eventi macroevolutivi non possono essere ricondotti nell'ambito della microevoluzione, e Simpson e Rensch hanno elaborato valide generalizzazioni riguardanti la macroevoluzione, senza dover analizzare mutamenti correlati nelle frequenze geniche. Questo atteggiamento è coerente con la definizione moderna di evoluzione, secondo cui l'evoluzione consiste in un mutamento di caratteristiche adattative e in una diversificazione, piuttosto che in un mutamento di frequenze geniche. La macroevoluzione è un campo di ricerca autonomo nell'ambito degli studi sull'evoluzione. Quel che sappiamo sulla macroevoluzione è frutto del lavoro dei paleontologi e dei tassonomisti. Le ricerche dei genetisti, effettuate all'interno delle popolazioni, consentono solo deboli inferenze circa i processi macroevolutivi, o per lo meno, per meglio dire, così è stato finora. Si spera che le analisi della struttura del DNA del genotipo, fatte dai biologi molecolari, porteranno alla fine a una comprensione della macroevoluzione di gran lunga più approfondita di quella raggiunta attraverso gli studi dei paleontologi e dei tassonomisti.
8. Come avviene il mutamento evolutivo.
La formula ‛l'evoluzione è un mutamento graduale di frequenze geniche nelle popolazioni', che per molto tempo è stata in auge negli scritti dei genetisti, è stata di recente messa in discussione dai naturalisti, che hanno avanzato una teoria alternativa, secondo cui gran parte dell'evoluzione è ‛spasmodica', nel senso che presenta improvvisi scatti evolutivi alternati a lunghi periodi di stasi (standstill). Per capire bene la natura di questo argomento, si deve rammentare che da Darwin in poi nessuno ha mai sostenuto che il tasso medio di mutamento evolutivo sia lo stesso in tutte le stirpi evolutive. In effetti è noto ai paleontologi da più di cent'anni che certi tipi, come il granchio reale (Limulus) o il gamberetto fatato (Triops), sono poco cambiati in più di cento milioni di anni, mentre in altri gruppi si sono verificati pronunciati mutamenti in meno di un milione di anni e, in qualche caso, persino in meno di centomila anni. Comunque la nuova teoria sul modo in cui avviene il mutamento evolutivo va al di là della semplice constatazione che diverse linee filetiche presentano diversi tassi di evoluzione.
9. La teoria degli equilibri di interpunzione.
Eldredge (1971) ed Eldredge e Gould (v., 1972) hanno proposto una teoria, la teoria degli equilibri di interpunzione, secondo cui tutti i più significativi mutamenti evolutivi si verificano durante brevi periodi di speciazione e le specie risultanti, dopo essersi affermate e aver proliferato, non cambiano sostanzialmente per milioni di anni e alla fine si estinguono. Questa teoria, pertanto, pone l'accento su due fatti indipendenti: a) la capacità innovativa della speciazione; b) la completa stasi delle specie pienamente evolute.
A sostegno di questa teoria esistono numerose prove empiriche. Ne indichiamo alcune.
1. Quando si esaminano le tracce di una specie fossile lungo la sequenza degli strati geologici, o non si riscontra alcun mutamento morfologico, oppure si riscontra un mutamento limitato a graduali variazioni di dimensioni o di proporzioni, che non corrispondono mai all'evoluzione di un nuovo taxon superiore o all'acquisizione di una vera e propria innovazione evolutiva.
2. Fra le specie viventi, le popolazioni più divergenti si trovano quasi esclusivamente in zone periferiche isolate, al di là dei confini del territorio continuo della specie, e si tratta di popolazioni che o hanno già completato la speciazione o si trovano nel corso di questo processo. In altre parole un mutamento morfologico significativo sembra essere associato all'origine di nuove specie.
Sulla base di queste osservazioni Mayr (v., 1954) ha avanzato l'ipotesi delle rivoluzioni genetiche. La forte diminuzione di variazione genetica in queste popolazioni fondatrici, combinata con l'inincrocio e l'omozigosi e con una selezione molto intensificata (nel nuovo ambiente fisico e biotico) della popolazione isolata, potrebbe portare a una ristrutturazione piuttosto drastica del genotipo e a un allentamento dei vincoli di sviluppo che controllano il fenotipo della popolazione parentale (rivoluzione genetica).
Questa ipotesi differisce per molti aspetti dalla teoria saltatoria di Goldschmidt (1940), secondo cui la macroevoluzione sarebbe determinata dalla nascita di mostri destinati ad affermarsi. In base a tale teoria si postulava che il mutamento evolutivo fosse dovuto alla nascita di un singolo individuo drasticamente diverso, che diventasse l'immediato progenitore di un tipo evolutivo completamente nuovo. Inoltre Goldschmidt postulava il verificarsi di mutazioni sistemiche, cioè di trasformazioni globali del genoma, mentre una tale possibilità è chiaramente incompatibile con la struttura del genotipo fondata sul DNA, come è stata decifrata dai genetisti molecolari.
Anche se esistono molte prove empiriche a favore della tesi che i maggiori progressi evolutivi e l'acquisizione di nuove caratteristiche evolutive si realizzino all'interno di poche popolazioni fondatrici durante il processo di speciazione, sussistono tuttora molte incertezze al riguardo. In particolare non si dispone ancora di alcun tipo di informazione circa la genetica della speciazione. Dobbiamo sperare che le ricerche in atto nel campo della genetica molecolare potranno permetterci di stabilire quale delle due teorie sia corretta, se quella secondo cui ogni mutamento evolutivo corrisponderebbe semplicemente a un'accelerazione del processo di ‛mutamento delle frequenze geniche', o quella secondo cui durante la fase di speciazione avrebbe luogo una ristrutturazione fondamentale del genotipo, avente come conseguenza un reindirizzamento di certi processi di sviluppo.
Ancor più controversa è la seconda tesi della teoria degli equilibri di interpunzione, quella di una completa stasi di tutte le nuove specie, una volta concluso il processo di speciazione. Attualmente sembra che si debba dare alla questione una risposta plurima. In effetti la maggior parte delle specie rinvenute nei reperti fossili mostrano di aver subito pochi cambiamenti dal momento della loro comparsa al momento della loro estinzione. D'altra parte sembra vi siano casi ben stabiliti, benché forse molto rari, di mutamento continuo di una linea evolutiva; comunque nessuno di questi casi, a quanto pare, dà luogo all'emergere di nuovi taxa superiori.
Quel che è più interessante, ovviamente, è la questione del perché si verifichi la stasi. In generale si ammette che essa sia dovuta al fatto che, una volta che si sia prodotto un apparato di sviluppo ben bilanciato, la selezione normalizzatrice rimuove tutti i devianti. Questa spiegazione, ovviamente, non rende conto del fatto che la selezione normalizzatrice interviene in tutte le linee evolutive e non impedisce rapidi mutamenti evolutivi in alcune di esse. Presumibilmente si dovranno distinguere due tipi di selezione stabilizzatrice, una riduzionista, che colpisce singoli geni e singole mutazioni aberranti, e una sistemica che colpisce componenti ben integrate del genotipo. Anche in questo caso solo la genetica molecolare potrà fornirci una risposta definitiva.
10. L'origine evolutiva di nuove strutture.
Uno dei quesiti preferiti che gli avversari di Darwin solevano porgli dopo la pubblicazione dell'Origine delle specie era: ‟Come può la selezione naturale render conto dell'origine apparentemente improvvisa di nuove strutture nella filogenesi, come i polmoni dei Vertebrati o le ali degli Insetti? Come potrebbe la selezione naturale costruire gradualmente queste strutture, specialmente nei casi in cui stadi intermedi, come ‛mezze ali', non possono in alcun modo offrire un qualche vantaggio selettivo?" Darwin faceva notare che l'acquisizione di nuove caratteristiche evolutive poteva dipendere dall'uno o dall'altro di due distinti processi: il primo processo consiste in un' ‛intensificazione di funzione' (A. N. Severcov), come nel caso della trasformazione delle estremità anteriori dei Mammiferi in ali (pipistrelli), pale (talpe), pinne (Cetacei), zampe (antilopi) o braccia (Primati). Nel caso dell'occhio, per esempio, Darwin fece notare, correttamente, che tutto ciò di cui c'era bisogno era una cellula epidermica fotosensibile, dopodiché la selezione avrebbe gradualmente costruito una retina e tutti gli organi accessori. Studi successivi, effettuati da anatomisti comparati e da microscopisti elettronici, hanno pienamente confermato l'ipotesi di Darwin, giacché ora è evidente che gli occhi hanno avuto origine almeno quaranta volte nel regno animale e si può quasi individuare un qualche stadio intermedio fra un'area sensibile alla luce sull'epidermide e i complessi occhi degli Artropodi, dei Cefalopodi e dei Vertebrati.
L'altro modo in cui possono avere origine nuove strutture evolutive è attraverso un mutamento di funzione. In base a questo principio un tale percorso per l'acquisizione di una nuova caratteristica evolutiva è disponibile ogniqualvolta un organo o una struttura può svolgere simultaneamente due funzioni. Ciò è vero per quel che riguarda le estremità anteriori dell'antenato degli Uccelli, che servivano sia per la locomozione sia per planare; o per quel che riguarda le antenne dei Crostacei Cladoceri, che funzionano sia come organi sensori sia come ‛pagaie'. Nel caso di un mutamento di funzione di questo tipo una struttura già completamente formata può mettersi al servizio di una nuova funzione. Le obiezioni dei critici di Darwin sono quindi state vittoriosamente confutate.
11. L'origine di taxa superiori.
I taxa superiori, sia dei Vertebrati (Mammiferi, Uccelli, Rettili), sia degli Insetti (farfalle, scarafaggi, cavallette), sia di qualsiasi altro phylum animale o vegetale, risultano sempre separati gli uni dagli altri da vistose lacune. La domanda che fu posta a Darwin fu: ‟Come si possono conciliare queste discontinuità con la teoria dell'evoluzione graduale?" Darwin rispose che dovevano essere esistiti degli anelli di connessione, che, però, dovevano essersi estinti da molto tempo. In effetti furono ben presto scoperti i cosiddetti ‛anelli mancanti': fra Uccelli e Rettili (Archaeopteryx), fra Mammiferi e Rettili (rettili con caratteristiche da mammiferi), fra Anfibi e Pesci (Ichthyostega) e fra altri gruppi di organismi. Molto spesso, comunque, gli stadi di connessione non sono presenti nei reperti fossili. Questo fatto può essere spiegato come segue: una qualsiasi nuova conquista di un'importante zona adattativa produce una reazione a catena di perfezionamenti e di estinzioni talmente rapida che ben presto fra il nuovo ‛tipo' e i suoi antenati si determina una netta discontinuità. Dato che gli anelli di connessione hanno durata molto breve e occupano un'area geografica limitata, è probabile che non si troveranno mai nei reperti fossili. Le lacune più grandi sono quelle che dividono i primi phyla di eucarioti del primo Cambriano e del tardo Precambriano. Vi sono buoni motivi per ritenere che i percorsi di sviluppo dei primi eucarioti fossero relativamente semplici e altamente flessibili, e che cambiamenti da un ‛tipo' a un altro fossero relativamente facili e rapidi. Una volta che i percorsi di sviluppo si furono stabilizzati, nel Cambriano, non si è più avuta l'evoluzione di un solo nuovo phylum di animali.
12. Progresso evolutivo.
Lamarck sosteneva che l'evoluzione portasse a una perfezione sempre maggiore. Questa idea concordava col concetto, proprio della teologia naturale, che la creazione divina dovesse essere perfetta. Darwin, che sulle prime nutriva idee del genere, presto le abbandonò, quando si rese conto dell'enormità del fenomeno dell'estinzione, della frequente presenza di organi rudimentali e di altre apparenti imperfezioni di natura organica. Egli giunse alla conclusione che la selezione naturale rende gli organismi perfetti quel tanto che basta perché possano competere, con successo, con altri organismi. Certo la selezione naturale è un processo di ottimizzazione, ma, come abbiamo detto prima, numerosi vincoli impediscono che si raggiunga la perfezione (ottimalità).
Quando si passa in rassegna la storia della vita a partire dai primi procarioti di 3,5 miliardi di anni fa fino alle Angiosperme, agli Insetti, agli Uccelli e ai Mammiferi attuali, non si può non riconoscere una complessità crescente. Questo processo di trasformazione spesso, e non senza giustificazione, è stato considerato un progresso evolutivo. Alcuni stadi di questo progresso sono: l'origine degli Eucarioti (presumibilmente dovuta alla simbiosi fra certi Procarioti), l'evoluzione di Metazoi e Metafiti con lo sviluppo di organi capaci di consentire una divisione del lavoro, la comparsa degli animali a sangue caldo, Uccelli e Mammiferi, lo sviluppo di un complesso sistema nervoso centrale e l'elaborazione di sofisticate cure parentali.
Pur ammettendo questa tendenza, il darwinismo non fa appello a cause finalistiche, ma sostiene che la variabilità genetica, la selezione e la competizione, combinate con l'estinzione, possano soddisfacentemente rendere conto della progressione della vita dalle sue forme primitive alle sue forme attuali.
13. L'evoluzione dell'Uomo.
Gli stadi attraverso cui l'Uomo si è evoluto a partire dai suoi antenati primati sono già stati descritti in un altro articolo di questa Enciclopedia (v. uomo: Origine ed evoluzione). Da un punto di vista del tutto generale, nell'ottica della biologia evolutiva, il progresso più notevole verificatosi nel corso dell'evoluzione dell'Uomo è stato lo sviluppo di un nuovo sistema di trasmissione dell'informazione, altamente elaborato. Nella maggior parte degli Invertebrati, in particolare negli Insetti, i genitori muoiono prima che i figli escano dall'uovo: tutta l'informazione comportamentale di un individuo è contenuta nel suo DNA. Soltanto nelle forme più evolute quest'informazione viene trasmessa anche attraverso un vero e proprio sistema di apprendimento: solo in quelle specie che manifestano cure parentali altamente sviluppate, come certi uccelli e certi mammiferi, i giovani hanno l'opportunità di apprendere dai genitori nonché da altri consanguinei e, in alcuni casi, da altri membri della specie. In questo modo l'informazione può essere trasmessa di generazione in generazione senza essere inclusa nel programma genetico.
Nei pochi animali in cui questo sistema di trasmissione dell'informazione non genetico esiste, esso è piuttosto rudimentale. Soltanto nell'Uomo il sistema di trasmissione culturale ha acquisito un'importanza fondamentale: anche prima dell'invenzione della scrittura e della stampa la tradizione orale svolse un ruolo decisivo nello sviluppo graduale della cultura umana e nell'esercitare la forte pressione selettiva che ha portato allo sviluppo di un cervello capace di immagazzinare e utilizzare questo tipo di informazione. Lo sviluppo del linguaggio, caratteristica esclusiva della specie umana, ha determinato, attraverso un certo numero di feedbacks positivi, la nascita e lo sviluppo della letteratura, della matematica, delle scienze e delle arti.
Questo sistema di trasmissione dell'informazione, accanto al riconoscimento dell'importanza dell'apprendimento, ha indotto alcuni studiosi a trascurare l'informazione innata dell'Uomo; ma recenti ricerche, condotte, in particolare, da psicologi infantili, hanno mostrato quanto sia ricco il repertorio di informazioni di cui l'Uomo nasce dotato. Nello studio dell'Uomo è della massima importanza tenere nella giusta considerazione entrambi i sistemi di trasmissione dell'informazione.
14. Il futuro della ricerca nel campo dell'evoluzione.
Malgrado le conoscenze raggiunte negli ultimi 125 anni, sarebbe prematuro pensare che ogni aspetto dell'evoluzione sia stato chiarito. Benché l'impalcatura teorica fondamentale del darwinismo sia ragionevolmente sicura, esistono ancora enormi vuoti nella comprensione di singoli fenomeni: non sappiamo ancora quanto siano importanti forme di speciazione diverse da quella geografica (allopatrica) e da quella per poliploidia; gli esatti dettagli dell'origine della vita restano ancora inspiegati; gli affini più stretti di alcuni importanti gruppi principali di animali e piante non sono ancora stati identificati; le ragioni della stasi evolutiva di tante specie costituiscono ancora un problema.
L'aspetto di gran lunga meno compreso del processo evolutivo è il ruolo svolto dalla struttura del genotipo: quale ruolo svolgono i mutamenti del genotipo durante la speciazione peripatrica rapida? Qual è il corrispettivo genotipico dei vincoli nello sviluppo? Qual è l'esatta natura dei cosiddetti geni regolatori e quanti tipi ne esistono? In che modo il comportamento agisce come forza selettiva? Quale relazione esiste fra speciazione e struttura di una popolazione, in particolare per quel che riguarda la tendenza a diffondersi?
Questa è solo una piccola cernita dei numerosi problemi, concernenti l'evoluzione, non ancora risolti. L'impostazione in termini evolutivi ha arricchito molto tutte le branche della biologia. Gran parte della biologia comportamentale e ambientale è attualmente impegnata in questioni evolutive, dato che nessun problema biologico può essere compreso appieno finché non si sia data risposta ai ‛perché'. Nessuna struttura, nessun comportamento, nessuna forma di adattamento, nessun tipo di distribuzione si può pienamente comprendere finché la relativa storia evolutiva non sia stata chiarita. È questo stato di cose che sancisce la validità dell'affermazione di Dobzhansky: ‟Nulla ha senso in biologia se non alla luce dell'evoluzione".
Bibliografia.
Alcock, J., Animal behaviour, Sunderland, Mass., 1979.
Barigozzi, C. (a cura di), Mechanisms of speciation, New York 1982.
Bendall, D. S. (a cura di), Evolution from molecules to men, Cambridge 1983.
Bock, W., Preadaptation and multiple evolutionary pathways, in ‟Evolution", 1959, XIII, pp. 194-211.
Bowler, P. J., The eclipse of Darwinism. Anti-Darwinian evolution theories in the decades around 1900, Baltimore 1983.
Brandon, R. N., Burian, R. M. (a cura di), Genes, organisms, populations, Cambridge, Mass., 1984.
Burkhardt, R. W. Jr., The spirit of system. Lamarck and evolutionary biology, Cambridge, Mass., 1977.
Campbell, B. (a cura di), Sexual selection and the descent of man, 1871-1971, Chicago 1972.
Darwin, Ch., On the origin of species by means of natural selection or the preservation of favored races in the struggle for life, London 1859.
Darwin, Ch., On the origin of species, (facsimile della prima edizione, a cura di E. Mayr), Cambridge, Mass., 1964.
Eldredge, N., Gould, S. J., Punctuated equilibria: an alternative to phyletic gradualism, in Models in paleobiology (a cura di T. J. M. Schopf), San Francisco 172, pp. 82-115.
Ford, E. B., Ecological genetics, London 1964.
Futuyma, D. J., Evolutionary biology, Sunderland, Mass., 1979.
Gould, S. J., Lewontin, R., The spandrels of San Marco and the Panglossian paradigm: critique of the adaptationist programme, in ‟Proceedings of the Royal Societ. London" (Series B), 1979, CCV, pp. 581-598.
Grene, M. (a cura di), Dimensions of Darwinism, Cambridge 1983.
Küppers, B.-O., Molecular theory of evolution, Berlin 1983.
Margulis, L., Origin of eukaryotic cells: evidence and research implications for a theory of the origin and evlution of microbial, plant and animal cells on the Precambrian earth, New Haven 1970.
Maynard Smith, J., The evolution of sex, London 1978.
Mayr, E., Change of genetic environment and evolution, in Evolution as a process (a cura di J. Huxley, A. C. Hardy, E. B. Ford), London 1954, pp. 157-180.
Mayr, E., Animal species and evolution, Cambridge, Mass., 1963.
Mayr, E., Behavior programs and evolutionary strategies, in ‟American science", 1974, LXII, pp. 650-659.
Mayr, E., Evolution and the diversity of life, Cambridge, Mass., 1976.
Mayr, E., Evolution, in ‟Scientific American", 1978, CCXXXIX, pp. 47-55.
Mayr, E., The growth of biological thought, Cambridge, Mass., 1982.
Mayr, E., How to carry out the adaptationist program?, in ‟American naturalist", 1983, CXXI, pp. 324-334.
Mayr, E., Provine, W. (a cura di), The evolutionary synthesis, Cambridge, Mass., 1980.
Mills, S. K., Beatty, J. H., The propensity interpreation of fitness, in ‟Philosophy of science", 1979, XLVI, pp. 263-286.
Ruse, M., The Darwinian revolution, Chicago 1979.
Schopf, T. J. M. (a cura di), Models in paleobiology, San Francisco 1972.
Sober, E., The nature of selection, Cambridge, Mass., 1984.
Stanley, S. M., Macroevolution. Pattern and process, San Francisco 1979.
Trivers, R., Social evolution, Menlo Park, Cal., 1985.
Wallace, B., Basic population genetics, New York 1981.
White, M. J. D., Animal citology and evolution, London 19733.
White, M. J. D., Modes of speciation, San Francisco 1978.
Williams, G. C., Adaptation and natural selection, Princeton 1966.
Wilson, E. O., Sociobiology, Cambridge, Mass., 1975.
Winne-Edwards, V. C., Animal dispersion in relation to social behaviour, Edinburgh - London 1962.
Evoluzione culturale di Luigi L. Cavalli-Sforza
SOMMARIO: 1. Definizioni di cultura. □ 2. La cultura tra gli animali. □ 3. Meccanismi della trasmissione culturale. □ 4. Cinetica dell'evoluzione culturale. □ 5. Fattori di evoluzione culturale: a) l'innovazione; b) la selezione; c) migrazione e deriva. □ 6. Distinzione fra trasmissione genetica e trasmissione culturale. □ 7. Esempi di trasmissione ed evoluzione culturale nell'Uomo. □ 8. Interazioni fra genetica e cultura. □ Bibliografia.
1. Definizioni di cultura.
Non vi è consenso sulla definizione di cultura. Già nel 1952 esistevano, in base a una ricerca di Kroeber e Kluckhohn (v., 1952), 164 definizioni diverse, classificabili in 6 categorie maggiori e 10 sottocategorie. Una così straordinaria varietà deriva soprattutto dal disaccordo, fra gli antropologi che amano autodefinirsi culturali, sui confini da dare alla disciplina. La proposta che seguiremo è quella di usare la definizione corrente in molte lingue indoeuropee, ed è stata fatta da Cavalli-Sforza e Feldman (v., 1981) in un libro che seguiremo anche in alcune altre sezioni di questo articolo. La definizione data nel Dizionario della lingua italiana di G. Devoto e G. C. Oli può servire da esempio: ‟Il complesso delle manifestazioni della vita materiale, sociale e spirituale di un popolo, in relazione alle varie fasi di un processo evolutivo o ai diversi periodi storici o alle condizioni ambientali". È una definizione molto vasta, il cui vantaggio principale non è, o non è soltanto, nell'evitare nuove parole o nuove formulazioni (specie ove esistono già numerosi precedenti), ma è concettuale e deriva dall'unità di fondo che lega tutti i fenomeni culturali, anche disparati fra loro come quelli della cultura materiale (utensili, tecnologia) e quelli della cultura sociale e spirituale. Quest'unità si manifesta soprattutto quando si studiano i meccanismi di variazione, trasmissione e conservazione: in pratica, quando i fenomeni culturali sono considerati sotto il profilo evolutivo. Se non vi fosse una ragione del genere, non avrebbe forse molto senso riunire sotto una stessa etichetta fenomeni diversi fra loro quanto quelli della vita spirituale e di quella materiale.
Il biologo, abituato a considerare i fatti della vita da un punto di vista evolutivo, non si sorprende di sentir dire che i fenomeni biologici sarebbero incomprensibili se non se ne studiasse l'evoluzione. Il pensiero evolutivo non ha avuto, in antropologia, uno sviluppo equivalente e lo studio dell'evoluzione culturale ha trovato pochi appassionati. Il più esplicito è stato L. White; molti peraltro hanno preferito limitare il significato dell'evoluzione culturale all'evoluzione dei sistemi politici, che naturalmente sono solo una parte della disciplina. Recentemente un certo numero di ricercatori, in buona parte ma non esclusivamente biologi, hanno più o meno indipendentemente scoperto che i fattori evolutivi che sono stati riconosciuti di importanza basilare nell'evoluzione biologica cioè mutazione, selezione, migrazione e deriva genetica ricompaiono, sotto altri nomi e con proprietà inevitabilmente diverse, nelle spiegazioni dell'evoluzione culturale. Dedicheremo a queste analogie alcuni capitoli di quest'articolo, avvertendo che esse sono utili soprattutto a livello di modelli, e che occorre tener presenti le differenze oltre che le somiglianze. Si deve riconoscere che questo modo di pensare è particolarmente utile a chi ha già famigliarità con i concetti dell'evoluzione biologica, ma senza dubbio le strutture concettuali di processi evolutivi anche diversi tra loro, come sono quelli dei geni e delle idee, hanno profonde somiglianze, che sono specialmente apprezzabili a un livello di astrazione adeguato. Un'applicazione che queste analogie hanno reso possibile è lo sviluppo di modelli matematici dell'evoluzione culturale, che hanno preso ispirazione da modelli creati per la biologia. Le differenze tra le due evoluzioni, peraltro, non consentono un trasporto diretto dei modelli biologici se non in pochi casi. Le teorie della biologia servono soprattutto come ispirazione generica; di solito è necessaria la creazione di modelli specifici per l'evoluzione culturale. Come in tutte le applicazioni matematiche alla biologia, questi modelli sono utili non tanto perché permettono di descrivere e prevedere con particolare esattezza i fenomeni studiati - anzi, in linea di massima, descrizione e previsione sono largamente approssimative - ma perché consentono di raggiungere delle generalizzazioni interessanti.
Negli ultimi decenni sono stati descritti comportamenti di animali che sono chiaramente di natura culturale. In genere la ‛cultura' può esistere ove siano presenti ambedue le capacità, di apprendere e di comunicare. È allora possibile generare e mantenere un'eredità culturale. La capacità di apprendere è largamente diffusa tra gli animali, ma naturalmente vi è una gamma grandissima di sofisticazione. I livelli più elevati si trovano solitamente, ma non sempre, nell'Uomo, ma comunque non mancano esempi di superiorità degli animali. Infatti non si può restare che pieni di ammirazione per le capacità di molti animali di compiere senza errori migrazioni lunghissime, ed è chiaro che, se mai abbiamo avuto queste straordinarie abilità, le abbiamo ora perdute. L'Uomo si distingue forse ancor più nettamente da tutti gli altri animali per quanto riguarda la capacità di comunicazione. Vi sono linguaggi naturali anche in uccelli e in altri mammiferi, ma di solito svolgono funzioni limitate e non hanno mai la ricchezza di struttura e di vocaboli caratteristica del linguaggio umano. Non vi è dubbio che abbiamo nel linguaggio la chiave per capire le profonde differenze a livello culturale tra Uomo e animali, anche quelli che gli sono più vicini. Siamo abituati a considerare l'Uomo come un animale unico fra tutti, ma molte osservazioni degli ultimi anni hanno diminuito la solidità del piedestallo su cui l'Uomo ha posto se stesso: non vi è dubbio tuttavia che il linguaggio umano è superiore di uno o più ordini di grandezza a quello degli animali per complessità e funzionalità, e che questa differenza di per sé può già spiegare almeno una buona parte delle differenze culturali tra Uomo e animali.
L'estrema cautela che alcuni ricercatori adottano nel parlare di cultura tra gli animali - ad esempio, alcuni preferiscono chiamarla ‛precultura', altri parlano di ‛tradizioni' per indicare particolari costumi trasmessi culturalmente negli animali - non sembra in realtà giustificata, né sembra necessario proteggere l'Uomo introducendo terminologie diverse per fenomeni affini negli uomini e negli animali. Le differenze tra Uomo e animali a livello dei fenomeni culturali sono abbastanza elevate da non lasciare dubbi sul primato culturale dell'Uomo.
2. La cultura tra gli animali.
Sono noti da parecchio tempo esempi di impiego di strumenti da parte di animali, ma si direbbe che solo con la scoperta che le scimmie antropomorfe, e in particolare gli scimpanzé, fanno uso di un certo numero di utensili semplici la comunità scientifica abbia accettato l'idea che la cultura materiale non è appannaggio esclusivo della nostra specie. È vero che la lista di strumenti diversi sembra essere particolarmente ricca per gli scimpanzé (sono stati descritti l'uso di rami e bastoni come armi, il lancio di pietre e di bastoni, l'uso di rametti e fili d'erba per ‛pescare' le termiti e quindi mangiarle, l'uso di foglie per bere e per pulirsi, e altri ancora) ma sono stati descritti anche molti altri esempi di impiego di utensili da parte di altri animali, e almeno alcuni meritano di essere qui ricordati. L'apertura di muscoli e ostriche viene praticata con tecniche diverse da diversi animali. La lontra di mare (Enhydra lutris), una rara specie di lontra marina del Pacifico settentrionale, usa galleggiare sulla schiena quando apre le conchiglie battendole contro una pietra che tiene sul petto e usa come incudine. In Europa il ratto ha recentemente appreso a usare i denti per aprire i molluschi che trova sul letto dei fiumi: i gusci aperti che si trovano in zone diverse mostrano differenze locali nella tecnica di apertura. La beccaccia di mare (Haematopus ostralegus) apre i gusci delle cozze con una di queste due tecniche: martellandoli col becco finché rompe il guscio, oppure, sott'acqua, introducendo il becco fra le valve e usandolo come lama per tagliare il muscolo che serra le valve. Probabilmente vi è una predisposizione genetica innata, ma il particolare metodo usato è appreso e non innato, come hanno dimostrato esperimenti di scambio di uova fra martellatori e tagliatori. Altri esempi descritti includono l'uso di pietruzze tenute tra le mandibole per chiudere l'entrata al nido (vespe solitarie del genere Ammophila), la rottura di uova col lancio di pietre (Hamirostra melanosterna, in Australia), e così via.
Alcune innovazioni e la loro diffusione sono state studiate in una ricerca di lunga durata su colonie di macachi giapponesi (Macaca fuscata). I risultati di alcuni esperimenti fatti dagli scienziati giapponesi su questi primati sono tra i più stimolanti di tutti gli studi di cultura tra gli animali, e in particolare la colonia dell'isola di Koshima fu protagonista delle osservazioni più interessanti. Gli studiosi cominciarono a lasciare sulla spiaggia patate dolci, intese come supplemento della dieta. I macachi dimostrarono interesse e accettarono il nuovo cibo. Dopo parecchi mesi fu notato un nuovo comportamento insorto spontaneamente: le patate venivano lavate nell'acqua prima di venir mangiate. L'invenzione fu opera di una femmina giovane di nome Imo, e si diffuse abbastanza rapidamente a tutte le scimmie più giovani di lei. Le scimmie più anziane mostrarono molta resistenza ad accettare la novità e solo alcune, tra le femmine più vecchie di Imo, acquisirono il comportamento. Le femmine che avevano imparato trasmisero l'abitudine ai propri figli.
Nella colonia avvennero altre innovazioni. L'abitudine di offrire i cibi sulla spiaggia, vicino al mare, portò molti macachi a prendere dimestichezza coll'acqua e col nuoto, e uno nuotò fino a un'altra isola. Seppellendo noccioline nella sabbia, in modo che l'operazione venisse notata dagli animali, si osservò che tutti gli individui di età non troppo vecchia né troppo giovane impararono a scavare per trovare le noccioline. Seminando chicchi di grano sulla sabbia, si osservò una nuova invenzione di Imo: i grani vennero raccolti e gettati in mare, dove i chicchi restarono a galla e la sabbia andò a fondo. Questa operazione di ‛flottazione' è simile a quella usata nell'industria mineraria per estrarre minerali da giacimenti alluvionali.
Un altro classico esempio di diffusione di una innovazione tra gli animali è quello delle cince. In Inghilterra divenne necessario cambiare i coperchi di stagnola delle bottiglie di latte che venivano lasciate sulle porte di casa poiché le cince avevano appreso a perforare il coperchio di stagnola col becco e a bere la crema. L'abitudine si diffuse rapidamente ai congeneri, per imitazione.
Questi studi ci mostrano sia innovazione che apprendimento di comportamenti che, almeno in parte, hanno un chiaro significato adattativo; il materiale non è sufficiente per dichiarare che vi sono grosse differenze individuali nella capacità di innovazione, ma la storia di Imo è certo molto suggestiva. Nella specie umana siamo abituati alla figura dell'inventore ed è anche ben noto che i singoli individui differiscono quanto a recettività all'innovazione. Molto interessante è la dimostrazione, nei macachi, che fattori di età e sesso, e forse di posizione sociale, hanno una profonda influenza sulla recettività.
La comunicazione tra gli animali è stata oggetto di studi numerosi. In genere, Vertebrati e Insetti (soprattutto gli insetti sociali) fanno uso di un certo numero di segnali diversi (da 10 a 40). Si tratta di atteggiamenti, movimenti o anche suoni; anche l'Uomo usa gesti e segnali mimici che costituiscono un vasto repertorio di comunicazione non verbale (100-200). Specie nell'ultimo decennio vi sono stati numerosi esperimenti per insegnare una comunicazione non verbale ai primati più vicini all'Uomo: col linguaggio dei sordomuti, con l'uso di tastiere di calcolatori speciali, di oggetti speciali, e così via. Questi studi hanno avuto un buon successo e hanno mostrato che i primati più vicini all'Uomo possono usare, per esprimersi, alcune centinaia di simboli, e anche costruire nuove associazioni. Non vi è accordo sulla loro capacità di dare struttura a frasi formate da parecchi simboli.
Dalle osservazioni della diffusione delle innovazioni si direbbe che l'imitazione semplice sia il meccanismo più comune per l'acquisizione di nuovi comportamenti, presenti in alcuni individui della popolazione, da parte di altri. Non vi è dubbio peraltro che i genitori, madre o padre o ambedue secondo le specie, possono talora svolgere un compito didattico attivo; ma purtroppo l'informazione al proposito è largamente aneddotica. Un'osservazione di particolare interesse è riportata in uno studio sull'alimentazione degli scimpanzè. È noto che gruppi diversi di scimpanzé hanno abitudini alimentari diverse, nutrendosi di un repertorio diverso di piante, pur se parecchie delle piante mangiate dalla colonia A e non dalla B sono presenti in ambedue gli ambienti. È stato osservato che una madre scimpanzé ha portato via dalla mano del figlio un frutto, che apparteneva a una specie ‛proibita', che il figlio si apprestava a mangiare.
I genitori non sono gli unici insegnanti, anche fra gli animali, pur se sono probabilmente di gran lunga i più importanti. Un gruppo africano di cercopitechi di savana usa cinque diversi segnali di allarme, a seconda del pericolo che si presenta, la comparsa di un'aquila, di un serpente, o di altri animali pericolosi e, a seguito dell'allarme dato da un individuo del gruppo, i singoli individui compiono azioni adeguate a controllare il pericolo. All'insegnamento del segnale partecipano non solo i genitori ma tutto il gruppo, che con le sue reazioni, o assenza di reazioni, mostra ai giovani cercopitechi se il segnale emesso da uno di essi è corretto o errato.
Lo studio della cultura animale ci mostra che l'adattamento culturale non compare improvvisamente nella specie umana, ma certo si sviluppa nella nostra specie più che in qualunque altra. Anche la semplice analisi delle localizzazioni funzionali sulla corteccia cerebrale ci mostra che fra l'Uomo e i cugini più prossimi vi è una notevole differenza nell'area di corteccia destinata a due funzioni motorie fondamentali che si esercitano nei processi culturali: il controllo della mano, che permette la creazione degli utensili ai quali è affidata la nostra cultura materiale, e il controllo della lingua, della bocca e degli altri organi responsabili della fonazione.
3. Meccanismi della trasmissione culturale.
La cultura si trasmette da una generazione all'altra; poichè solo una piccola parte potrebbe essere creata di nuovo in una generazione, la trasmissione delle conoscenze accumulate nelle generazioni precedenti permette l'accrescimento continuo nel tempo del nostro patrimonio culturale. La scrittura e gli altri mezzi di registrazione dell'informazione di cui oggi disponiamo permettono un accumulo molto più efficiente di quanto sarebbe possibile con la sola tradizione orale.
La trasmissione della cultura è quindi una parte integrante del fenomeno culturale, e il suo studio sistematico una necessità che è stata finora assai poco sentita. Cominceremo con il separare e classificare i diversi meccanismi di trasmissione. Un interessante risultato dello studio di questi meccanismi è la possibilità di prevedere la cinetica dell'evoluzione culturale e la variazione fra individui e fra gruppi.
Possono esservi diverse fasi nella trasmissione: di solito sono almeno due, e sono state sistematicamente studiate in alcuni casi di diffusione dell'innovazione. La prima fase è l'apprendimento dell'esistenza di una innovazione, che può avvenire per stadi diversi: possiamo chiamarla la fase della comunicazione. L'innovazione non viene di solito accettata automaticamente, ma solo una percentuale, magari piccola, di coloro che hanno ricevuto l'informazione reagisce all'inizio positivamente; Rogers e Shoemaker (v., 1971) ritengono che alcuni individui o gruppi sono predisposti ad accettare più facilmente l'innovazione, altri invece sono più tardivi. Abbiamo visto che almeno un dato di osservazione dei fenomeni culturali in animali superiori è perfettamente in accordo con questa interpretazione. Rogers e Shoemaker ritengono che la variazione del tempo di latenza individuale (cioè del tempo di accettazione) sia la causa principale se non unica della lentezza con la quale certe innovazioni si diffondono nella popolazione. Vi sono peraltro circostanze in cui i tempi della comunicazione e la relativa variabilità individuale sono i primi responsabili.
Esistono situazioni in cui le due fasi, comunicazione e accettazione, non sono separabili. Un esempio estremo è quello dell'imprinting, per cui un individuo esposto a uno stimolo adeguato durante un particolare periodo della sua vita (il periodo critico o sensibile) ne viene influenzato in modo indelebile e il suo comportamento è determinato in modo irreversibile per il resto della vita dagli eventi di quel periodo. L'esempio classico è l'identificazione della madre in oche, anatre e altri uccelli. Il primo oggetto che si impone all'attenzione del pulcino entro 24 ore dalla nascita viene identificato come la madre, anche se è un essere vivente di un'altra specie o addirittura un oggetto inanimato, purché mobile. In queste situazioni l'accettazione è immancabile se l'esposizione avviene nel periodo sensibile. Un altro esempio: esperienze su animali mostrano che le preferenze di cibo sono influenzate dall'alimentazione della madre durante l'allattamento. Non è ben noto se esistono nell'Uomo situazioni estreme come quella che ha dato origine al concetto di imprinting, ma certo vi sono molte esperienze, soprattutto della vita infantile o giovanile, di cui sappiamo, pur se solo su base aneddotica, che hanno una profonda influenza nel seguito della vita. Manca un loro studio sistematico.
In genere, comunque, l'accettazione della maggioranza dei suggerimenti o insegnamenti che ci vengono dati ha ampi margini, e così la selezione culturale può spesso esercitarsi con discreta, e talora ampia, libertà.
Vi sono almeno quattro categorie di meccanismi della trasmissione culturale. Naturalmente è raro che essi agiscano in completo isolamento, ma di solito la trasmissione avviene in più di un modo. La prima categoria che considereremo, la trasmissione da genitore a figlio, è comune tra gli animali e certamente anche nell'Uomo. La chiameremo, in accordo con la definizione usata dagli epidemiologi, ‛verticale'. Può accadere che uno o ambedue i genitori partecipino alla trasmissione (trasmissione uniparentale o biparentale); la forma uniparentale è soprattutto frequente quando un carattere culturale è limitato a uno dei sessi. La trasmissione verticale è di importanza particolare durante la giovane età, che è anche quella maggiormente sensibile e formativa. Nella società moderna molti compiti didattici sono assolti dalla scuola e vi sono ampie occasioni per subire l'influenza dei coetanei: di conseguenza l'importanza culturale dei genitori è in diminuzione e i meccanismi di trasmissione sono in continua modificazione.
I contatti con i coetanei, o anche con persone di un'altra generazione al di fuori dei genitori, si possono descrivere come affini, formalmente, a quelli che consentono la diffusione delle malattie infettive, nel senso che possono seguire gli stessi modelli concettuali e matematici. Si deve, però, dare naturalmente un significato profondamente diverso alle costanti che intervengono nella diffusione dei fatti culturali e in quella delle malattie infettive. I modelli relativi si possono perciò chiamare ‛epidemici', e formano una seconda importante categoria di meccanismi di trasmissione, cui si può dare il nome di ‛orizzontale', sempre seguendo l'uso epidemiologico. Si è preferito però distinguere, nel caso culturale, la situazione in cui la diffusione avviene fra individui dello stesso gruppo d'età, o più semplicemente della stessa generazione, che chiameremo trasmissione ‛orizzontale' propriamente detta, da quella della diffusione tra un individuo della generazione precedente e uno della generazione successiva che non siano in rapporto di genitore e figlio, che chiameremo trasmissione ‛obliqua'. La distinzione fra trasmissione obliqua e orizzontale è utile perché esse hanno cinetiche diverse, e anche quella tra obliqua e verticale, perché nella verticale solo un individuo (o al massimo due), quello che ha il ruolo sociale di genitore, può avere funzione di trasmettitore nei confronti di un individuo della generazione successiva, mentre in quella obliqua sono molti i trasmettitori in potenza. Si può anche aggiungere che la trasmissione fra individui di generazioni diverse, e quindi di età diverse, è di solito nella direzione dal più vecchio al più giovane, ma la regola non è assoluta.
Nella trasmissione orizzontale si ha un'equazione molto semplice per descrivere la modificazione del fenomeno in due intervalli di tempo successivi. Se p è la frequenza di individui portatori del carattere in un certo momento, e dopo il passaggio di un'unità di tempo (di lunghezza arbitraria) la frequenza di individui diventa p′, l'equazione più semplice che si può scrivere è:
p′ = p + cp (1 − p), (1)
ove c è una costante che si può chiamare coefficiente di trasmissione, o di ‛conversione' di individui che non portano il carattere in individui che lo portano, nell'unità di tempo scelta.
Questa equazione definisce una curva molto nota, detta logistica, che quando l'unità di tempo scelta è sufficientemente piccola va da un valore iniziale, al tempo zero, fino al 100% che viene raggiunto asintoticamente (a tempo infinito), con una velocità determinata da c. Le curve ottenibili modificando e complicando il processo mantengono solitamente un andamento sigmoide da un minimo a un massimo, che può essere inferiore al 100%, ma in certe condizioni possono acquistare un andamento oscillante, che si può effettivamente osservare sia nelle malattie infettive che nei fenomeni culturali. La trasmissione orizzontale può essere limitata a gruppi e sottogruppi sociali, gruppi di età, e così via.
La trasmissione verticale da sola non determina necessariamente la diffusione del carattere all'intera popolazione. Nella sua forma più semplice, quella uniparentale, essa è descritta dall'equazione
p′ = bp, (2)
ove b, il coefficiente di trasmissione verticale, è la probabilità che il figlio di un genitore portatore del carattere porti anch'esso il carattere per effetto della trasmissione verticale. L'unità di tempo che separa p, la frequenza del carattere nei genitori, da p′, la frequenza del carattere nei figli, è una generazione. In un esempio numerico che daremo nel capitolo successivo considereremo la trasmissione biparentale con coefficienti qualunque, e quattro tipi possibili di incrocio fra due genitori con e senza il carattere.
Nella trasmissione verticale uniparentale descritta dalla (2) la frequenza del carattere non può salire oltre il valore iniziale, ma anzi scende a zero se b è minore di uno. Poiché b è definito come una probabilità, non può essere superiore a uno; se è uguale a uno, la frequenza del carattere nella popolazione resta costante nel tempo. Il coefficiente b è uguale a uno se i figli di un genitore portatore del carattere mostrano sempre il carattere: è una condizione limite che verrà raramente raggiunta. Naturalmente, se coesistono altre forme di trasmissione, esse possono portare la frequenza del carattere a qualunque valore.
Se da una parte la trasmissione verticale ricorda quella biologica, che pure avviene da genitori a figli, e quella orizzontale in senso lato ricorda la trasmissione delle malattie infettive , vi sono due altri meccanismi che sono invece caratteristici della sola trasmissione culturale. In uno dei due vi è un solo trasmettitore con molti riceventi: è la situazione tipica dell'insegnamento scolastico, con l'insegnante da una parte, gli allievi dall'altra. La stessa situazione si ritrova nelle società umane meno primitive quando si hanno strutture politiche con capi e sudditi: l'informazione trasmessa è allora più spesso paragonabile a un ordine. Non solo nelle strutture politiche, ma anche in quelle religiose, si ha di solito un'autorità riconosciuta che detta il comportamento dei sudditi o dei credenti. Ordini e informazioni sono diffusi da una fonte centrale a un gruppo che può essere numerosissimo. La complessità dei gruppi sociali spesso richiede l'organizzazione di gerarchie, in cui il rapporto di capo e dipendenti si ripete a ogni gradino della gerarchia. La struttura gerarchica del gruppo consente a una persona sola di estendere la propria influenza a un gruppo grandissimo, attraverso molti gradini intermedi. Peraltro nel mondo moderno i mass media e i mezzi elettronici diffondono l'informazione e influenzano l'opinione pubblica secondo lo schema di trasmissione culturale che stiamo considerando, ma senza far uso di intermediari e in un tempo brevissimo. Chiameremo questi meccanismi di trasmissione ‛un trasmettitore-molti riceventi', o anche uno-a-molti'.
L'altro meccanismo tipico della trasmissione culturale è l'esatto opposto del precedente: molti trasmettitori influenzano un individuo solo ('molti trasmettitori-un ricevente' o ‛molti-a-uno'). Di solito, in questo caso, gli stessi trasmettitori influenzano anche gli altri individui del gruppo, in modo solitamente concorde. Questo modo di trasmissione è particolarmente efficace, in quanto ogni ricevente subisce l'influenza e la pressione di molti individui, magari di tutti gli altri membri del gruppo, compresi i più anziani e più influenti, con risultati quasi irresistibili. Chiamiamo questa trasmissione anche col nome di ‛pressione del gruppo sociale'. Nella fig. 1 abbiamo rappresentato graficamente i quattro tipi di trasmissione culturale secondo la classificazione da noi proposta. Vedremo nel seguito che i quattro tipi hanno conseguenze diverse agli effetti della cinetica dell'evoluzione culturale e anche della variabilità che ci si può attendere tra individui di un gruppo e tra gruppi.
4. Cinetica dell'evoluzione culturale.
Abbiamo rappresentato il processo elementare dei fenomeni culturali come l'acquisizione di un carattere culturale da parte di un individuo di una popolazione. Il particolare carattere culturale considerato può essere uno qualunque: l'affiliazione a una religione, l'accettazione di un nuovo dogma, l'apprendimento di una lingua o di una nuova parola, l'acquisto di un oggetto, l'apprendimento di una tecnica nuova, il voto a un candidato, un'opinione, un costume, un'abitudine, una credenza particolare. La descrizione di un fenomeno più complesso sarà inevitabilmente più complicata. Difficoltà maggiori si hanno, naturalmente, nella descrizione di caratteri che cambiano nel corso della vita di un individuo più volte e in parecchie direzioni. In genere, ogni descrizione concreta richiede definizioni accurate del carattere in esame, ed è più soddisfacente se si scelgono caratteri che possono essere rilevati con l'opportuna riproducibilità.
I modelli che verranno esposti si applicano con maggior facilità a caratteri semplici, cioè a quelli che una volta acquisiti non si modificano nel corso della vita. Nulla vieta naturalmente di complicare i modelli per adattarli alle esigenze particolari delle situazioni concrete, ma è più facile raggiungere conclusioni convincenti con modelli semplici di fenomeni semplici. Le generalizzazioni che descriveremo sono il frutto della ricerca teorica di Cavalli-Sforza e Feldman e valgono indipendentemente dal tipo di carattere considerato. Le due equazioni date nelle formule (1) e (2) sono gli esempi più semplici dei modelli di trasmissione ed evoluzione culturale che si applicano con meccanismi di trasmissione di tipo orizzontale e verticale, rispettivamente. Complicando ed estendendo il meccanismo di trasmissione si ottengono, naturalmente, equazioni più complesse, che spesso non hanno una soluzione se non per via numerica. Ma le tecniche di analisi matematica dei processi evolutivi consentono di raggiungere almeno certe conclusioni, per esempio sugli stati finali del processo, sulla sua velocità e sull'influenza delle variabili di maggior importanza.
È opportuno notare il rapporto che esiste tra la trasmissione e l'evoluzione. La trasmissione è il passaggio di informazione da un individuo a un altro, ma il modo in cui abbiamo definito le costanti di trasmissione nei due esempi indicati sopra include inevitabilmente anche la seconda fase, che segue alla comunicazione, e cioè l'accettazione. La trasmissione è quindi il processo elementare completo. Se consideriamo insieme, per via statistica, tutti i processi elementari di trasmissione che avvengono nell'intera popolazione per un determinato carattere, abbiamo descritto l'evoluzione della popolazione in un certo intervallo di tempo. Le due equazioni riportate sopra si riferiscono all'intervallo più breve in cui è utile considerare il processo. Nel caso della trasmissione verticale l'intervallo di tempo è di una generazione, poiché descriviamo il passaggio dell'informazione da genitore a figlio, e perciò è relativamente lungo se riferito alla durata della vita di un individuo, ma pur sempre corto se riferito a millenni di storia culturale. Per fare previsioni su tempi più lunghi di una generazione bisogna avere conoscenze maggiori: ad esempio, dei valori di b in generazioni diverse. Queste non sono di solito disponibili, ed è facile ottenerle solo per le generazioni viventi. Previsioni sono ancora possibili ammettendo che il valore di b rimanga costante nelle generazioni, ma questo spesso non è vero, e i cambiamenti di b nel tempo sono particolarmente importanti oggi che l'influenza dei genitori sui figli va diminuendo. Talora è possibile valutare almeno grossolanamente le conseguenze di una variazione nel tempo delle costanti che influenzano il processo. In alcuni casi potrà trattarsi di una variazione nel tempo, in altri potranno esservi variazioni frequenza-dipendenti.
Nel caso della trasmissione orizzontale l'unità di tempo è meno chiaramente definita che nella trasmissione verticale e viene scelta in modo arbitrario, ma è in genere assai più piccola di una generazione. La formula (1) è una ricorrenza, che si risolve con i metodi delle equazioni alle differenze finite; per intervalli di tempo piccolissimi si passa all'uso di equazioni differenziali, che danno risultati di solito più semplici ma molto simili, almeno per valori non troppo estremi delle costanti in esame.
La soluzione delle ricorrenze (1) e (2) porta alle conclusioni che abbiamo già indicato, in forma molto elementare e grossolana, nel capitolo precedente. Di solito alla trasmissione verticale partecipano ambedue i genitori, generando formule più complesse della (2); spesso si trovano presenti anche altri meccanismi di trasmissione.
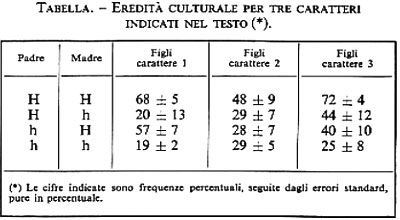
Come esempio citiamo una ricerca su un gruppo di studenti americani, in cui si studiarono le relazioni tra genitori e figli per un certo numero di caratteri. La tabella ne mostra un estratto: in essa il carattere 1 è la frequenza della preghiera; il carattere 2 la fiducia nell'importanza dell'abilità personale nel determinare il proprio destino (in opposizione all'opinione che solo la fortuna conta); il carattere 3 le simpatie politiche verso sinistra. Si è indicato con H un valore elevato del carattere (cioè al di sopra di una soglia arbitraria che è di solito quella corrispondente alla mediana del carattere) e con h un valore piccolo, cioè al di sotto della soglia. Esaminando un certo numero di famiglie, scelte per convenienza come triadi padre-madre-figlio/a, si sono trovati i risultati della tabella.
Nel seguito ammetteremo che la frequenza del carattere nei figli sia espressione della sola trasmissione culturale verticale, ma questa ipotesi è assai limitativa. Per esempio, si ignorano gli effetti dello stato socioeconomico, che pure è trasmesso da genitori a figli e potrebbe avere effetti talora simili, ma anche più complessi, e si ignora la possibilità di trasmissione biologica. Il primo caso si può controllare con altre tecniche statistiche, il secondo è più difficile da controllare (v. cap. 5, È c).
La semplice ispezione dei dati mostra che nel carattere 1 (frequenza della preghiera) la madre sembra avere un'influenza importante, mentre il padre non ne ha alcuna (trasmissione uniparentale). Questo fatto da solo depone a favore della trasmissione culturale, dato che sono eccezionalmente rari i meccanismi di trasmissione biologica uniparentale. Nel carattere 2 è solo l'azione concertata dei due genitori che sembra avere influenza, cioè assistiamo a una trasmissione biparentale con effetto sinergico. Nel carattere 3 ambedue i genitori contribuiscono con influenza pressochè eguale e le loro azioni sembrano sommarsi (trasmissione biparentale additiva). In tutti e tre i casi, comunque, anche in assenza dell'influenza dei genitori (cioè quando ambedue i genitori sono di tipo h) il carattere si può ritrovare nei figli, pur se con frequenza inferiore. Ciò può indicare che giocano anche altri meccanismi di trasmissione.
Una cautela di rigore è ispirata alla possibile esistenza di effetti dell'età sulla frequenza del carattere. Gli studenti che hanno fornito i dati della tabella sono stati esaminati a un'età diversa da quella dei loro genitori, e per alcuni caratteri vi sono variazioni sistematiche con l'età.
Ignorando per il momento questa possibile difficoltà, che richiede dati di altro genere, si può calcolare la frequenza che il carattere prenderà ‛all'equilibrio', cioè dopo il passaggio di molte generazioni, se la trasmissione verticale, e quelle dovute ad altri meccanismi che pure contribuiscono, rimangono di intensità costante nel tempo. Per questo calcolo ha importanza anche l'eventuale correlazione per il carattere tra padre e madre, che può essere determinata sia dalla scelta matrimoniale (se non direttamente per il carattere in esame almeno per altri correlati) sia dall'influenza reciproca dei due genitori nel corso della loro vita comune. I dati riportati sinteticamente nella tabella non sono utili per valutare questa correlazione, che però non è trascurabile negli esempi indicati. Ciò malgrado, la struttura dei coefficienti di trasmissione nei dati della tabella, soprattutto per i caratteri 1 e 3, fa sì che non vi sia una grande influenza della correlazione fra genitori sull'evoluzione del carattere. Chiamando b3 il coefficiente di trasmissione (frequenza nei figli) per le coppie di genitori H × H, b2 per le coppie H × h, b1 per le coppie h × H, b0 per le coppie h × h, alla fine del processo si ha una frequenza pa (valore asintotico di p) data da
pa = b0/(1 − b1 − b2 + 2 b0), (3)
se b3 + b0 = b1 + b2, e una formula lievemente più complicata se b3 + b0 è diverso da b1 + b2. Negli esempi della tabella per il carattere 1 la frequenza attesa all'equilibrio (il valore asintotico) è 32,9%, mentre quella dei genitori è 58,7%, e quella dei figli 50%. La stima della frequenza del carattere all'equilibrio può essere non valida se le ipotesi sulle quali è basata non si verificano, per esempio se l'effetto di trasmissioni diverse da quella verticale, che è riassunto nel coefficiente b0, non è costante nel tempo, e così via. Per il carattere 2 il valore asintotico è 30,3% contro 34,6% nei genitori e 31,3% nei figli. Nell'ultimo carattere il valore di pa è 41,5% contro 75,4% nei genitori e 59,2% nei figli, una discesa quindi anche maggiore che per il primo carattere. Molti caratteri mostrano questo comportamento, probabilmente dovuto al fatto che l'influenza dei genitori è discesa negli ultimi decenni.
In genere, la trasmissione verticale assicura alta stabilità a un sistema culturale, poiché l'evoluzione sotto di essa è particolarmente lenta. Un motivo molto semplice è che l'unità di tempo in cui un cambiamento può avvenire, la generazione, è particolarmente lunga, dell'ordine di 25-30 anni. Inoltre, questo meccanismo di trasmissione è simile alla trasmissione biologica, che è il meccanismo più conservatore che esista. Difatti, sotto trasmissione biologica semplice non vi è alcuna evoluzione: la frequenza di un carattere rimane costante indefinitamente e le misure medie e di variazione di un carattere quantitativo non mostrano alcun cambiamento nel tempo. Ciò è vero anche nella trasmissione culturale uniparentale se il valore di b nell'eq. (2) è eguale a uno, o in una situazione biparentale come quella indicata nella tabella, quando l'equilibrio è stato raggiunto o quando i quattro coefficienti b0, b1, b2, b3 acquistano certi valori semplici.
È facile intendere che nella trasmissione orizzontale od obliqua la velocità di evoluzione può essere elevatissima, perché l'unità di tempo nel processo (1) può essere molto breve e il valore della costante c elevato. Nel caso elementare illustrato dalla (1) il valore asintotico è il 100%, ma il carattere non si diffonde necessariamente a tutta la popolazione, sia perché possono esservi stratificazioni con barriere efficaci alla comunicazione fra strati, sia perché altri fattori possono intervenire a modificare il fenomeno. Spesso vi è trasmissione verticale congiunta ad altre, per esempio orizzontale od obliqua, e lo studio degli effetti congiunti mostra gli equilibri possibili.
Gli altri due tipi di trasmissione, che abbiamo indicato col nome di uno-a-molti o, viceversa, di molti-a-uno, si possono considerare come varianti della trasmissione orizzontale od obliqua in cui compaiono effetti speciali, che sono stati studiati matematicamente in dettaglio. È peraltro facile prevedere quel che ci si può attendere in situazioni limite. La trasmissione uno-a-molti, ovvero maestro-allievi o capo-dipendenti, rende massima la caratteristica fondamentale della trasmissione obliqua od orizzontale: l'elevata velocità di trasmissione, che qui diventa quasi infinita. La trasmissione molti-a-uno, o pressione del gruppo sociale, si può anche vedere come una variante della trasmissione obliqua od orizzontale, nel senso che ogni trasmettitore agisce su più ricevitori (in senso concorde). Anche qui la velocità iniziale di evoluzione è elevata; il risultato, l'omogeneizzazione completa del gruppo sociale, è raggiunto rapidamente in quanto tutti propinano a tutti lo stesso insegnamento (qualunque sia il carattere in questione), e in breve tempo il gruppo è omogeneo.
Ciò indica un altro aspetto dell'evoluzione culturale che merita di essere studiato in dettaglio, e cioè la variazione tra individui del gruppo e l'effetto su di essa di un particolare meccanismo di trasmissione. Accanto a questa variazione ('entro i gruppi') è importante considerare anche quella ‛tra gruppi', intendendo per gruppi popolazioni separate da barriere fisiche o sociali sufficienti a ridurre o eliminare completamente lo scambio culturale e l'intermigrazione di individui. In linea di massima, la variazione tra gruppi ('sincronica', cioè osservata simultaneamente in gruppi geograficamente diversi) è molto simile alla variazione ‛diacronica', cioè quella dello stesso gruppo in tempi diversi. La velocità di variazione diacronica è naturalmente molto simile, come concetto, alla velocità di evoluzione. Lo studio della variazione si compie particolarmente bene per caratteri quantitativi, in cui si separano i concetti di valore medio e di variabilità di un carattere. Nel caso di caratteri qualitativi, come negli esempi finora considerati (che erano in realtà caratteri quantitativi ridotti a qualitativi per semplicità, introducendo una soglia arbitaria), media e variabilità sono confuse l'una con l'altra, in quanto la percentuale di individui portatori del carattere esprime il valore medio della popolazione e anche la sua variabilità.
La trasmissione verticale tende a conservare, anche nel caso culturale, la variabilità esistente nella popolazione, tranne che in situazioni biparentali per caratteri quantitativi, in cui la media viene conservata ma la variabilità entro il gruppo scende a un valore minimo. La trasmissione orizzontale od obliqua tende a eliminare la variabilità e i due meccanismi di uno-a-molti e molti-a-uno sono estremi in questo senso: ambedue tendono a creare una totale omogeneità del gruppo sociale per il carattere in esame. Naturalmente l'effetto di omogeneizzazione nel caso della trasmissione uno-a-molti è limitato al gruppo in cui un dato capo politico, religioso o spirituale, o un particolare insegnante agisce. Quando si osserva una situazione di totale conformità non vi è quindi bisogno di invocare il conformismo, inteso come tendenza più o meno innata a comportarsi come gli altri, poiché i meccanismi di trasmissione culturale caratteristici della società umana moderna (uno-a molti) e anche quelli che si trovano pure in società tradizionali (molti-a-uno) hanno un estremo effetto omogeneizzante.
Vi è una differenza importante nella velocità con cui una innovazione può essere accettata da un gruppo quando viene proposta da un individuo. Sotto trasmissione verticale la diffusione all'intera popolazione è lentissima e procede con velocità paragonabile a quella con cui un mutante biologico viene fissato in una popolazione: migliaia, decine di migliaia di anni o anche più, secondo le dimensioni della popolazione. Sotto trasmissione orizzontale può essere veloce, anche se parte del gruppo può restare sul momento non influenzato o influenzabile. Sotto trasmissione di uno-a-molti l'accettazione è immediata e universale (o quasi, sempre nel gruppo in cui la trasmissione avviene) e si ha quindi la massima velocità di accettazione. Nella trasmissione di molti-a-uno vi è la massima resistenza all'accettazione e quindi, dopo l'iniziale omogeneizzazione, la velocità diviene praticamente nulla. Il conservativismo di certi gruppi sociali trova probabilmente spiegazione nel meccanismo di trasmissione verticale e/o di molti-a-uno con cui l'informazione solitamente si diffonde in questi gruppi: i cacciatori-raccoglitori e gli agricoltori hanno mostrato nei millenni un'estrema conservazione culturale, forse anche a causa dei meccanismi di trasmissione più comuni tra questi gruppi sociali. I caratteri culturali che hanno dimostrato la più alta conservazione sono quelli che si riferiscono ai costumi famigliari, che vengono naturalmente trasmessi entro l'ambito della famiglia con trasmissione verticale e pressione del gruppo famigliare.
La velocità dell'evoluzione culturale dipende anche da molti altri fattori. Un fattore forse importante, che non è stato adeguatamente studiato, è la maggiore sensibilità di alcune età al processo di trasmissione. Esistono probabilmente periodi critici durante i quali l'apprendimento di certi fatti culturali avviene con particolare facilità: le età infantili e giovanili sono probabilmente le più importanti, ma non le sole, in cui questo può avvenire. Le impressioni acquisite in periodi critici o sensibili possono essere largamente irreversibili. È interessante riflettere sul fatto che probabilmente questi periodi sensibili o critici rappresentano adattamenti biologici specifici.
Esamineremo con qualche dettaglio nel prossimo capitolo i fattori di evoluzione culturale di cui si può prevedere l'esistenza in base ad analogie, anche se superficiali, coi processi dell'evoluzione biologica.
5. Fattori di evoluzione culturale.
a) L'innovazione.
L'inizio di ogni cambiamento culturale è un'innovazione o invenzione, che avviene per opera di un individuo, spesso con un'intenzione precisa. L'innovazione può diffondersi al resto della popolazione (o a una parte della stessa), rapidamente o lentamente secondo il meccanismo di trasmissione e la natura dell'innovazione. Tranne che nel caso della trasmissione di uno-a-molti, si ha un aumento graduale della frequenza con cui il nuovo carattere, che è il risultato dell'innovazione, si estende alla popolazione o a parte di essa. Formalmente si può dire che allo stesso modo si ha l'insorgenza di una mutazione biologica in un solo individuo e la sua possibile diffusione al resto della popolazione; ma nel caso della mutazione biologica la diffusione avviene solo ai discendenti del primo portatore e ai discendenti dei discendenti, e quindi richiede un tempo molto lungo per la ‛fissazione', cioè per l'estensione al resto della popolazione. Allo stesso modo una novità culturale trasmessa esclusivamente per via verticale può richiedere un tempo altrettanto lungo; ordinariamente, peraltro, funzionano anche meccanismi diversi da quello verticale, che permettono una trasmissione culturale più rapida e, nel mondo moderno, rapidissima.
Per alcuni versi l'innovazione somiglia alla mutazione, anche se naturalmente nella sostanza si tratta di fenomeni diversissimi: la mutazione biologica è un cambiamento casuale in una struttura molecolare ben precisa, che è ereditario poiché la struttura affetta dalla mutazione è il modello che viene copiato nelle generazioni successive. Invece, le strutture biologiche coinvolte nell'innovazione sono del tutto ignote, in quanto si tratta di processi che si svolgono nel nostro cervello, la cui natura sfugge largamente alle nostre conoscenze attuali. Malgrado questa nostra ignoranza di base, è chiaro che vi sono differenze notevolissime tra la mutazione e l'innovazione, così come ve ne sono in genere fra le strutture coinvolte nell'evoluzione biologica e in quella culturale. Esistono però alcune somiglianze di forma, che rendono i due processi evolutivi trattabili con modelli matematici molto simili. Per continuare con il confronto tra biologia e cultura, aggiungeremo che la mutazione è rara e avviene in modo casuale; l'innovazione non è necessariamente rara ed è, almeno nella nostra percezione, un fatto di solito intenzionale, designato a ovviare a un inconveniente, pericolo o necessità, o a soddisfare un desiderio. In realtà vi è un elemento casuale anche nel cambiamento culturale, in quanto alcune innovazioni sono in pratica il frutto di errori involontari. Tipicamente, almeno una parte delle variazioni che si osservano nei manoscritti di testi tramandati dai copisti medievali è strettamente analoga a mutazioni casuali; altre sembrano variazioni intenzionali del copista. Indipendentemente dall'origine, volontaria o casuale, dei cambiamenti osservati nei manoscritti medievali, gli studiosi di testi antichi hanno potuto ricostruire la storia e le origini di vari manoscritti classici, usando tecniche molto simili a quelle usate per l'evoluzione molecolare. Un altro motivo, forse più importante, che parzialmente invalida l'intenzionalità di certe ‛mutazioni' culturali, è che l'intenzione dell'autore di un'innovazione e di coloro che l'accettano è spesso diversa dalle conseguenze reali dell'innovazione medesima. Per esempio, la motivazione espressa da coloro che praticano una certa norma può non avere alcun rapporto con le conseguenze derivanti dalla sua applicazione. Non vi è alcuna certezza che la circoncisione maschile sia benefica sul piano igienico o in altri modi noti. I motivi avanzati per giustificare la circoncisione femminile sono assai diversi secondo le popolazioni: ad esempio, l'eliminazione dell'adulterio attraverso l'eliminazione del piacere della donna, o il desiderio di concentrare l'attenzione della donna sul piacere dell'uomo, evitando la distrazione che può derivare dall'orgasmo. Molte popolazioni africane praticano un lungo tabù sessuale post partum anche di tre anni. La giustificazione avanzata da un gruppo che ha un tabù particolarmente lungo è che il rapporto sessuale interromperebbe l'allattamento, incidendo così gravemente sulla salute del neonato, che normalmente viene, in questi gruppi etnici, allattato per tre anni. In tutti i casi citati è probabile che le conseguenze maggiori siano di natura diversa da quella percepita, e che in genere le motivazioni avanzate dai sostenitori, e forse anche dagli iniziatori, siano di scarsa importanza rispetto alle conseguenze reali. L'importanza maggiore della circoncisione maschile può derivare dalle occasioni, cui dà origine, di tenere feste di notevole importanza sociale; quella femminile permette di riaffermare il controllo dell'uomo sulla donna, pur se a costo di infezioni e perdita della fertilità; un lungo tabù sessuale dopo il parto ha conseguenze importanti sulla natalità ed evita uno sviluppo demografico inopportuno, contribuendo all'equilibrio tra nascite e morti. La mancanza di rapporti tra i moventi percepiti e le conseguenze effettive di una norma rende l'intenzionalità dell'innovazione un fatto meno centrale di quanto potrebbe a prima vista apparire.
Vi sono alcune altre somiglianze tra la mutazione e l'innovazione, che vale la pena di ricordare. In genere si osserva che le innovazioni più importanti sono avvenute indipendentemente più di una volta, in luoghi diversi ma in tempi simili. Ciò è vero non solo per l'invenzione dell'automobile, della radio e del telefono in tempi assai vicini a noi, ma anche, per es., per l'invenzione dell'agricoltura o della ceramica in tempi assai più remoti. Naturalmente, più lontano è il tempo in cui queste innovazioni sono avvenute, più difficile è ricostruirne la storia o studiarne i dettagli e stabilirne l'indipendenza. Ciò era vero fino a poco tempo fa anche per la mutazione biologica. Un esempio è l'origine della falcemia, un'anemia ereditaria che, nei genitori degli anemici, assicura una maggior resistenza alla malaria, e che è molto diffusa in regioni malariche dell'Africa, Arabia e India. Fino a due anni fa non era noto se la falcemia fosse dovuta a una sola mutazione diffusa dal suo luogo di origine a una vasta regione, grazie al suo vantaggio in presenza di malaria, o se invece fosse stata originata molte volte indipendentemente. Le mutazioni sono rare, e la falcemia in particolare è dovuta alla sostituzione di un nucleotide con un altro in una sola posizione del gene dell'emoglobina; ci attendiamo che la sua frequenza sia molto bassa, dell'ordine di una cellula mutata su cento o mille milioni di cellule riproduttive. Le moderne tecniche di genetica molecolare hanno però recentemente permesso di analizzare la regione cromosomica vicino alla mutazione e di concludere in base ai risultati di quest'analisi che vi sono state molte origini indipendenti della mutazione in diverse aree geografiche. Simili conclusioni sono state raggiunte per un'altra malattia ereditaria che ha una diffusione simile alla falcemia, la talassemia. Analogamente, l'aumento delle conoscenze archeologiche potrà forse un giorno chiarire in modo inequivoco se le origini plurime dell'agricoltura o della ceramica furono davvero indipendenti.
L'invenzione di grosse novità tecnologiche è naturalmente un fenomeno raro, e inoltre dipende dagli sviluppi tecnologici precedenti. Per quanto grande fosse il desiderio dell'uomo di aumentare la propria mobilità, l'effettivo uso di meccanismi automobili ha dovuto attendere lo sviluppo di motori adatti. Finché il motore a benzina non è stato sufficientemente perfezionato si è trovato in concorrenza con il motore elettrico, che era ancora assai diffuso all'inizio del secolo. Date le difficoltà di costruire motori efficienti, il cavallo è rimasto per sei o settemila anni senza veri concorrenti. Ma non tutte le invenzioni sono così difficili; altre sono abbastanza semplici, tanto che, se una tecnica non ci viene insegnata direttamente, abbiamo buone probabilità di svilupparla da soli. Vi sono molti esempi della vita quotidiana in cui chiaramente la probabilità di innovazione è elevata.
b) La selezione.
La mutazione biologica e l'innovazione non sono la sola analogia tra evoluzione genetica ed evoluzione culturale. In realtà, tutti i fattori di evoluzione biologica - la selezione, la migrazione, la deriva casuale (drift) - hanno analoghi culturali. La somiglianza può essere anche più sostanziosa per questi fattori di quanto non lo sia per la mutazione. Vi sono però possibilità di equivoco che bisogna sottolineare. Tra i sostenitori dell'importanza della selezione nell'evoluzione culturale ricordiamo l'antropologo D. Campbell; riportiamo nel seguito però il contributo dato da Cavalli-Sforza e Feldman, i quali hanno dato particolare rilievo alla distinzione tra selezione culturale e selezione naturale. Per selezione culturale si intende la decisione in favore di un'innovazione, o contro di essa, la scelta tra alternative disponibili, e così via. È una scelta compiuta dall'individuo o da qualcun altro per lui. Purtroppo, le nostre conoscenze sui meccanismi che intervengono nelle nostre scelte e decisioni sono assai modeste. Possiamo tuttavia indicare fattori importanti, come l'intensità e la ripetizione dello stimolo, l'influenza di chi propone la scelta, la percezione dei vantaggi che ne risultano, le difficoltà possibili generate dall'accettazione, ma non abbiamo elementi per scendere alla base del processo psicologico, per non parlare di quello neurologico.
Talora una scelta vera e propria manca, in quanto la fase della decisione è assente, come nelle situazioni di imprinting, o quando la scelta vera è effettuata da un capo e non dai singoli individui che ne dipendono, e la disciplina è assoluta. Ma solitamente almeno una parte della decisione è affidata a noi; è chiaro che le difficoltà di comprensione di questi fenomeni al livello psicologico e anche a quello filosofico (il ‛libero arbitrio') sono tuttora grandissime. Non vi sono molte speranze di giungere rapidamente alla comprensione delle strutture e delle funzioni dei meccanismi biologici coinvolti. Anche per questo motivo la ricerca e la definizione di ‛unità' della trasmissione e dell'eredità culturali sembrano totalmente premature. I tentativi compiuti - per esempio la definizione di cultgen, secondo Lumsden e Wilson (v., 1981), o di meme secondo Dawkins - non hanno avuto molto successo. Mendel parlava di ‟elementi" ove oggi parliamo di geni, e in pratica si accontentava di osservare caratteri (che oggi chiamiamo ‛fenotipici') scelti ragionevolmente in modo che fossero netti e facili da classificare. Sarebbe difficile, oggi, far di meglio a livello di eredità culturale, e conviene scegliere caratteri arbitrari che siano facilmente osservabili.
Nella trattazione teorica della trasmissione culturale data sopra, la fase della decisione o scelta è incorporata nel meccanismo della trasmissione culturale e nei coefficienti di trasmissione, di cui fa parte integrante. Quindi gli studi di cinetica di cui abbiamo visto qualche esempio elementare includono la selezione culturale, ma non quella naturale, che va considerata come fattore aggiuntivo. Come è noto, per valutare l'intensità della selezione naturale si misura la fitness darwiniana di un certo tipo in esame, confrontandone il potenziale contributo alla generazione successiva con quello degli altri tipi esistenti nella popolazione in esame. Analogamente, per selezione culturale intendiamo qui la modificazione del numero di figli attesi di un individuo che ha adottato un certo carattere culturale, a confronto con individui che non hanno adottato quel carattere.
La fitness è composta di due variabili demografiche, la capacità di sopravvivere e quella di riprodursi, e ambedue possono influenzare il numero di figli. La componente di mortalità è assai meno importante nelle società umane più avanzate, in quanto le mortalità in età preriproduttiva e riproduttiva, che maggiormente contano agli effetti della fitness, sono quasi nulle. Vi è un gran numero di norme trasmesse culturalmente igieniche, mediche e di comportamento quotidiano - che permettono di mantenere elevata la probabilità di sopravvivere a lungo. È interessante però ricordare che vi sono anche condizioni di origine culturale, come ad esempio l'abuso di droghe, che agiscono in senso opposto e costituiscono indubbiamente un grave pericolo anche per la vita. Alcuni sport possono anche, seppur più raramente, mettere in pericolo la vita. In condizioni di guerra la mortalità è elevata e possono esservi differenze individuali di comportamento che modificano la probabilità individuale di sopravvivenza, con effetti di selezione naturale.
Agli effetti della selezione naturale, nelle società umane più sviluppate, la variazione individuale della fecondità è assai più importante di quella della mortalità. In quasi tutte le società umane, infatti, avanzate o meno, la fecondità è un fatto culturale. Le difficoltà che si hanno nel modificare la natalità dei paesi in via di sviluppo derivano dall'estrema resistenza al cambiamento di costumi che hanno spesso millenni di vita e che sono tra i meglio conservati, venendo instillati (probabilmente) al livello della famiglia, quindi con trasmissione verticale e con forte pressione dell'intero gruppo famigliare. È assai interessante notare, a questo proposito, che l'introduzione dei metodi di contraccezione è un classico esempio di fenomeno in cui la selezione culturale è in aperto contrasto con quella naturale. In una società in cui un gruppo segue la fertilità naturale (o, più esattamente, ha costumi che conferiscono una più elevata natalità) e un altro ha una natalità più bassa perché pratica un controllo delle nascite più elevato, alla lunga sarà sempre ‛vincente' il gruppo a più alta natalità, a meno che il controllo delle nascite più rigoroso non venga esteso, attraverso una continua campagna culturale, fino all'eliminazione dei costumi che comportano una maggiore natalità. Abbiamo già notato che l'uso di droghe è un chiarissimo esempio di totale contrasto fra la selezione culturale (la droga è di uso piacevole, e quindi la selezione culturale è a suo favore se non intervengono considerazioni a lunga scadenza) e la selezione naturale, che è invece contraria. Altri esempi di dissociazione fra selezione culturale e selezione naturale meritano di essere ricordati. Alcune malattie da virus vengono trasmesse ad alta frequenza per mezzo di costumi favoriti per ragioni culturali, e hanno certamente l'effetto di aumentare la mortalità. Un esempio è il kuru, che veniva trasmesso nella tribù dei Fore (Nuova Guinea) dalle pratiche cannibalistiche sui morti, specie sui genitori, fino a diffondersi a gran parte della tribù. Un altro esempio è l'epatite, che viene trasmessa soprattutto da madre a figlio con meccanismi di ordine non biologico, ma di comportamento; sono state di fatto escluse la trasmissione del virus attraverso la placenta e quella attraverso il latte, a favore di altre non ancora ben identificate, che avvengono probabilmente per via culturale e ambientale. Anche le sindromi di deficienza immunologica acquisita sono trasmesse in prevalenza da individui con attività omo- o eterosessuale abnormemente elevata. Queste attività eccezionali sono rese possibili da situazioni culturali particolari, ed è negli ambienti in cui queste situazioni sono più frequenti che la malattia raggiunge la sua massima incidenza.
Abbiamo insistito sulle discordanze tra selezione naturale e culturale perché ve ne sono di importanti e non sono poche. Di solito, però, vi è accordo fra le due selezioni, fatto abbastanza ovvio. Per esempio, molti degli insegnagnamenti dei genitori e della scuola sono volti a diminuire i pericoli e a consentire uno sviluppo fisico e spirituale armonico e adeguato all'ambiente fisico e sociale. Ma val la pena di insistere sul fatto, già notato, che la percezione dell'utilità delle regole trasmesse culturalmente non è sempre corretta. Le superstizioni sono per definizione esempi di atti di solito inutili, talora controproducenti e crudeli. Nella migliore delle ipotesi, l'utilità di un comportamento superstizioso non è mai stata provata, e non vi sono motivi razionali per credervi. Si direbbe che siano preda di superstizioni soprattutto persone che hanno subito una specie di imprinting (in senso lato), probabilmente in giovane età, dal quale non riescono a liberarsi anche perché la superstizione comporta un ricatto (la iettatura magari non esiste, ma se poi fosse vera?). Si può rompere una lancia in favore della superstizione affermando che vi è un vantaggio in pratiche superstiziose come per esempio l'astrologia (che ha una diffusione enorme anche tra le persone colte), nel senso che queste pratiche aiutano a superare situazioni di insicurezza e di ansia. Non si spiegano così, però, le enormi differenze di importanza della superstizione fra paesi e fra gruppi sociali. Si potrebbe dire che alcune superstizioni sono nate per diminuire la probabilità di eventi controproducenti: vi sono buone ragioni per evitare la perdita di sale e olio, specie quando sono particolarmente preziosi. È chiaro d'altra parte che il rimedio suggerito dalla superstizione, lo scongiuro di rito, non ha funzione deterrente, ma può servire solo a eliminare il timore superstizioso.
Un'altra ipotesi che spiega il successo di alcune pratiche magiche sembra interessante. È stato suggerito che la pratica della divinazione, com'è noto molto diffusa tra Egizi, Greci, Etruschi e Romani, corrisponda a quanto la teoria dei giochi dimostra conveniente in certe condizioni: quando non vi sono elementi per la scelta fra diverse strategie, conviene sceglierne una a caso. Un altro motivo importante è che lasciando ‛agli dei', cioè in pratica al caso, la scelta di una strategia si possono evitare conflitti tra coloro che sostengono strategie diverse; tali conflitti sono più probabili quando la scelta è difficile, e quindi tanto vale, in situazioni del genere, lasciarla al caso. Naturalmente, la convinzione che la decisione suggerita dagli dei è giusta può avere forza di persuasione sufficiente solo se si ha grande fiducia nella capacità di determinati individui, gli àuguri, di comunicare con la divinità, cioè se non vi è il sospetto che si tratta in realtà di una turlupinazione.
Avendo citato molti esempi in cui le nostre credenze trasmesse culturalmente sono di scarsa o nulla utilità, se non addirittura controproducenti, e se utili, lo sono per ragioni molto diverse da quelle effettivamente credute, vale la pena di citare qualche altro esempio in cui la selezione culturale è di diretto aiuto a quella naturale, cioè le due sono perfettamente parallele, o quasi. I proverbi, il cui insieme costituisce un ‛libro della saggezza' diffuso per tradizione orale, sono una buona testimonianza della validità di molta cultura popolare. Se consideriamo un altro aspetto interessante della cultura popolare, erbe e trattamenti curativi, troviamo senz'altro molti rimedi validi. Ma ve ne sono altri in cui è difficile avere una fiducia se non, al più, marginale, come la cura delle ferite con ragnatele, un'idea che alcuni cercano di difendere con la possibile (ma assai poco probabile) presenza di penicillina nelle ragnatele.
c) Migrazione e deriva.
Nell'analogia tra evoluzione biologica ed evoluzione culturale anche la migrazione e la deriva (drift) trovano interessanti estensioni. Il termine ‛migrazione' verrà usato, nel contesto culturale, per definire la comunicazione tra gruppi. Essa può essere strettamente culturale, nel senso che non coinvolge lo scambio fisico di individui, ma solo lo scambio di informazione; la comunicazione a distanza, che oggi è la più comune, fino a pochi anni fa era rara e sempre mediata da individui che si spostavano da un luogo all'altro: anche se non si trattava di una loro migrazione permanente, vi era sempre alla base della ‛migrazione culturale' un episodio di ‛migrazione biologica'.
Lo scambio culturale tra gruppi è il meccanismo che consente a gruppi diversi di non divergere all'infinito. Quando la differenza culturale comincia a essere elevata, la comunicazione diviene più difficile e frammentaria e la velocità di divergenza può aumentare. Un importante esempio (e causa) di divergenza culturale è la divergenza linguistica che, come è noto, evolve rapidamente verso l'incomprensibilità reciproca di linguaggi che si differenziano indipendentemente l'uno dall'altro, pur partendo da un unico progenitore. L'ordine di tempo dello sviluppo della incomprensibilità reciproca è di mille anni, e quindi relativamente breve. L'evoluzione linguistica è un soggetto di notevole interesse sia per se stessa sia come modello dell'evoluzione culturale. Val la pena di aggiungere che, nell'evoluzione linguistica, la migrazione culturale, come sopra è stata definita, viene indicata col nome di ‛prestito' (borrowing).
La migrazione di individui e idee da un gruppo all'altro, anche quando vi sia incomprensibilità delle lingue, consente di diminuire le differenze culturali e di trasmettere dall'uno all'altro gruppo innovazioni tecniche e di altra natura. Assistiamo nel mondo a un aumento continuo dello scambio di individui. Malgrado ciò, le culture più forti sopravvivono, se non intatte, almeno scarsamente modificate. Anche fisicamente, mentre in certe nazioni i diversi gruppi etnici mostrano mescolanze sempre più grandi, in altre vi è una maggiore resistenza all'incrocio. Malgrado gli scambi genetici, i sistemi culturali possono mantenersi più o meno intatti, grazie alle forze di conservazione presenti nella cultura. I fenomeni culturali possono mostrare variazioni rapide, ma anche gradi elevatissimi di stabilità.
La migrazione di grossi gruppi di individui ha conseguenze culturali importanti. Spesso si può avere semplicemente il trapianto in nuove regioni dei sistemi di vita praticati nel paese d'origine, ed è ciò che avviene regolarmente nella colonizzazione: gli abitanti dei paesi occupati dai coloni si trovano così a contatto con una società di solito più avanzata economicamente e ne ricevono influenze importanti (non sempre positive). L'influenza culturale dei secondi sui primi è di solito modesta. Invece, quando grossi gruppi da paesi più poveri migrano in altri economicamente più avanzati, si ha l'effetto opposto: pochi dei costumi originali rimangono e, se i figli degli emigrati nati nel paese di emigrazione vi si stabiliscono in modo permanente, si ha un'acculturazione elevata e relativamente rapida, che peraltro può richiedere varie generazioni per divenire totale.
La ‛deriva' (in inglese drift) è un termine che useremo qui con il significato che ha in genetica: variazione casuale delle frequenze di un carattere (gene nell'esempio biologico) attraverso le generazioni, dovuta al fatto che da una generazione all'altra variano, per ragioni casuali, le frequenze dei caratteri negli individui che effettivamente fungono da genitori. L'esempio più semplice di una situazione che promuove una deriva genetica elevata si ha quando vi sono pochi ‛fondatori' di una nuova popolazione, una colonia, un'isola, ecc. In questo caso i geni delle generazioni successive sono interamente determinati da quelli dei fondatori (salvo future mutazioni o immigrazioni); alcuni geni potranno essere molto rari o addirittura assenti tra i fondatori (e quindi fra i loro discendenti), per ragioni puramente casuali, tanto più facilmente quanto meno numerosi sono i fondatori; per altri geni avverrà l'opposto. Il calcolo delle probabilità permette di rendere quantitative queste considerazioni. L'effetto dei fondatori non è limitato alla prima generazione, ma avviene a ogni generazione, in quanto gli individui che si riproducono a un dato tempo sono i fondatori delle generazioni successive e solo da loro dipende quali geni saranno passati ai discendenti. L'effetto della deriva nella differenziazione genetica delle popolazioni umane è assai più importante di quanto si ritenesse una volta, ma naturalmente non è l'unica causa di differenziazione. Si deve anche notare che in linguistica la parola drift ha acquisito un significato diverso, indicando le tendenze, spesso tutt'altro che casuali, osservate in certi cambiamenti linguistici, che sovente hanno andamenti paralleli in lingue e tempi diversi (Shapir). Il parallelismo mostra che non si può trattare di deriva casuale; la casualità è invece sempre implicita nell'uso genetico del termine.
L'importanza della deriva casuale nell'evoluzione culturale è forse anche più grande che nell'evoluzione biologica. Infatti, non appena le società umane sono divenute più numerose, in pratica con l'avvento dell'agricoltura, i gruppi sociali sono divenuti più grandi, con l'inevitabile formazione di capi e di gerarchie, per cui la trasmissione uno-a-molti ha preso enorme importanza. L'influenza dei capi politici e spirituali si proietta nei costumi anche molto al di là di quella che è l'influenza politica o spirituale diretta. La moda dei vestiti o dei gusti e anche l'arte sono state spesso influenzate dalle scelte dei re o di individui influenti a corte. Con il declino delle monarchie la posizione di modello da imitare è stata presa da personalità popolari per altri motivi. Queste tendenze a imitare alcune persone in vista comportano rapidi cambiamenti nei gusti e persino nei costumi di larghe masse di individui, tutti attenti alle scelte di un solo ‛idolo'. Ne risultano cambiamenti rapidi e massicci, in quanto le preferenze di un solo individuo influenzano quelle di molti altri.
6. Distinzione fra trasmissione genetica e trasmissione culturale.
Il problema di distinguere, in un caso specifico, se un determinato carattere scelto per l'indagine sia trasmesso per via biologica o culturale può essere di assai difficile soluzione. Forse l'esempio più clamoroso è stato quello del quoziente di intelligenza (Q. I.), che è stato (ed è tuttora, in alcuni ambienti) considerato come quasi interamente determinato dall'eredità biologica di un individuo, mentre in altri ambienti scientifici viene considerato di determinazione puramente ambientale, cioè culturale. Il Q.I. è un modello che si presta bene per discutere i problemi della distinzione tra la trasmissione genetica e quella culturale.
Nell'analisi del Q.I. si trovano elevate correlazioni tra genitori e figli, e tra fratelli o sorelle, che depongono a favore dell'eredità biologica; la decisione in favore del modello biologico può sembrare ancor più inevitabile quando si aggiunga che è estremamente elevata la correlazione tra gemelli identici (cioè che hanno un corredo genetico identico), a differenza di quelli fraterni che sono geneticamente simili quanto due fratelli qualunque. Non vi è dubbio però che l'ambiente in cui crescono i figli, soprattutto quello più formativo per i giovani, è influenzato in modo importante dai genitori e che i fattori ambientali che contribuiscono in modo essenziale al Q.I. oltre la famiglia, e cioè la scuola e le amicizie, sono largamente determinati anch'essi in via indiretta dalla famiglia e soprattutto dal suo stato socioeconomico. Ciò potrebbe largamente o anche interamente spiegare le correlazioni tra genitori e figli, o tra fratelli. L'altissima correlazione tra gemelli potrebbe essere dovuta all'estrema somiglianza dell'ambiente in cui si trovano i gemelli identici, che normalmente vanno alla stessa scuola, hanno gli stessi amici e rimangono in stretto contatto per tutta la vita. Allo stesso modo, il fatto che la correlazione tra gemelli identici è superiore a quella tra gemelli fraterni non è sufficiente a convincerci che la causa della differenza sia biologica, perché la somiglianza di ambiente dei gemelli identici è più grande di quella dei gemelli fraterni.
Molti ricercatori per risolvere il problema hanno esteso gli studi ai bambini adottati in famiglie diverse da quella di origine, e ne hanno confrontato il Q.I. con quello dei genitori e parenti biologici, da una parte, e dei genitori e parenti adottivi dall'altra. Le correlazioni con i parenti biologici e adottivi dovrebbero dirci, rispettivamente, l'importanza della trasmissione biologica (cioè genetica) e di quella culturale. In linea di massima si trova correlazione con ambedue. Allo stesso modo, se si prendono gemelli identici cresciuti in famiglie diverse (che non sono peraltro sempre tanto isolate fra loro quanto per un buon ‛esperimento' sarebbe desiderabile) si trova una correlazione più bassa che tra gemelli cresciuti insieme.
Questi studi non sono completamente soddisfacenti per parecchie ragioni. Ad esempio, solo adozioni precocissime dovrebbero contare; inoltre, le agenzie di adozione collocano i bambini da adottare in famiglie che hanno somiglianze fisiche o socioeconomiche con quelle di origine, e possono quindi creare correlazioni spurie; infine i dati validi sulle adozioni sono scarsi. Comunque, le migliori analisi compiute recentemente sull'insieme dei dati disponibili danno all'incirca eguale importanza all'eredità culturale e a quella genetica. Le stesse difficoltà che per il Q.I. si incontrano per molti altri caratteri.
Quando la trasmissione è interamente o quasi di origine culturale è più facile giungere a una conclusione. In una analisi compiuta su un certo numero di caratteri della vita quotidiana che misurano abitudini, costumi, credenze ecc., come gli esempi riportati nella tabella, si sono trovati importanti effetti culturali, soprattutto nelle scelte e nelle abitudini religiose e in quelle politiche, con un effetto preponderante della trasmissione verticale. È stato possibile escludere quasi completamente effetti genetici, poiché per questi caratteri non si trova una differenza importante tra le correlazioni di gemelli identici e di gemelli fraterni. Quando ciò avviene sembra difficile poter sostenere che vi è un effetto genetico; bisognerebbe mostrare che c'è, ma è mascherato da altri fattori. Invece quando si osserva una correlazione tra gemelli identici maggiore che tra quelli fraterni, l'ipotesi dei fattori genetici è verosimile, ma è difficile escludere la possibilità che gli ambienti dei gemelli identici siano più simili di quelli dei gemelli fraterni, per ragioni esterne ai gemelli stessi. Quindi non si può escludere l'effetto culturale a favore della trasmissione biologica sulla base di soli dati sui gemelli, in assenza di adozioni.
7. Esempi di trasmissione ed evoluzione culturale nell'Uomo.
Nella nostra specie abbiamo una grande ricchezza ma allo stesso tempo povertà di esempi di evoluzione culturale: infatti quasi tutte le manifestazioni o attività umane, dalla tecnologia alle arti, alle scienze e alla storia, costituirebbero dei possibili esempi, ma sono state raramente studiate sotto il profilo evolutivo. Pochissimi casi sono stati analizzati quantitativamente, e ancora meno sotto il profilo dei meccanismi di trasmissione presenti, anche perché lo studio sistematico della trasmissione è molto recente. Se consideriamo che una caratteristica fondamentale della cultura è la possibilità di trasmetterla e accumularla, risulta evidente che lo studio della trasmissione dovrebbe essere una parte integrante dell'analisi dei fenomeni culturali.
Dei quattro meccanismi principali di trasmissione viene percepito di solito come il più caratteristico quello in cui il fenomeno elementare di trasmissione avviene attraverso il contatto tra due persone non apparentate. Come abbiamo già detto, esso ha una somiglianza formale con i meccanismi che danno origine alle epidemie ed endemie, in quanto la diffusione avviene attraverso il ‛contagio' di un individuo suscettibile da parte di un individuo infetto. Questo meccanismo di contagio è la base più comune dei fenomeni di ‛diffusione', parola usata da antropologi e archeologi per definire soprattutto il passaggio di novità culturali da un gruppo all'altro. Lo studio quantitativo della diffusione delle innovazioni è naturalmente molto più facile nelle culture occidentali moderne che in quelle antiche o tradizionali. Uno dei prototipi di maggior successo di analisi della diffusione nel mondo moderno è stato lo studio della diffusione geografica di innovazioni ad opera del geografo Hagerstrand (v., 1967), mentre la cinetica della diffusione delle innovazioni e i fattori di ordine non geografico che la influenzano sono stati analizzati da Rogers e Shoemaker (v., 1971). Lo studio degli effetti della propaganda nell'adozione di un qualunque prodotto commerciale è un esempio di analisi della diffusione, e nelle ricerche di mercato se ne incontrano molti. Peraltro la propaganda diretta all'acquirente è un caso di trasmissione uno-a-molti se viene fatta attraverso i mass media. Spesso si ha una trasmissione di questo tipo congiunta ad altre forme di propaganda che, realizzandosi attraverso il contatto personale con il possibile cliente, sono più vicine alla trasmissione per contagio. Una parte delle vendite, poi, avviene perché l'acquirente di un oggetto, se ne è soddisfatto, convince un suo conoscente all'acquisto; questa è forse la forma più tipica di contagio. Lo studio quantitativo mostra che, come nelle epidemie, la cinetica della diffusione delle innovazioni è di solito una curva di tipo logistico, come abbiamo già visto sopra, complicata talora dalla presenza di altre forme di trasmissione.
Abbiamo visto che il contagio, da solo o insieme con altri meccanismi di diffusione, è la forma di trasmissione culturale solitamente responsabile di cambiamenti culturali caratteristici perché rapidi (anche se meno rapidi di quelli ottenibili con la trasmissione uno-a-molti, che è una specie di contagio generalizzato e simultaneo). Anche se queste possono sembrare le due forme di diffusione culturale più caratteristiche, non si deve sottovalutare l'importanza degli altri meccanismi. Studi quantitativi hanno mostrato che la trasmissione verticale è particolarmente importante per alcuni tipi di caratteri, e continua a esserlo anche oggi, pur se nella vita moderna altri meccanismi tendono a prevalere. Esiste una ricchissima letteratura sull'influenza dei famigliari e in particolare del padre e della madre sul comportamento, ma purtroppo ben poche ricerche sono state condotte in modo da prestarsi a un'analisi quantitativa soddisfacente. La raccolta e il riassunto dei dati nella forma usata nella tabella permettono di verificare facilmente se le osservazioni si prestano a essere rappresentate con modelli semplici. Il modello additivo più semplice è il seguente:
xf = a + b1xp + b2xm,
dove xf è la frequenza con cui un carattere si manifesta nel figlio, xp e xm le frequenze nel padre e nella madre rispettivamente, e b1, b2 sono le probabilità o coefficienti di trasmissione paterno e materno, cioè numeri compresi nell'intervallo tra 0 e 1, che misurano la rispettiva importanza del padre e della madre. L'equazione è costruita sulla base di un modello secondo cui gli effetti del padre e della madre si sommano, e a essi se ne aggiunge un altro o altri esterni, misurati dalla costante a. Nei tre esempi della tabella questo modello è utile solo per i caratteri 1 e 3, ma in una serie più estesa di casi si è trovato che esso è quasi sempre valido. Quando non è valido si può utilmente sostituirlo con modelli più complessi, noti agli statistici.
Analizzando studenti americani si è trovato che alcuni gruppi di caratteri mostrano una trasmissione verticale particolarmente elevata. Si tratta di quelli connessi con la religione e con la politica. La fig. 2 mostra alcuni risultati della ricerca. La religione cui appartiene il figlio è, nei matrimoni misti osservati in questa ricerca, determinata quasi interamente dalla madre: lo mostra il fatto che il coefficiente materno è quasi pari a uno, mentre quello paterno è praticamente nullo. Così è per la frequenza della preghiera, mentre la frequenza della partecipazione alle funzioni religiose è determinata egualmente da ambedue i genitori. La trasmissione verticale è responsabile, secondo queste osservazioni, della quasi totalità del comportamento religioso, dato che la somma dei due coefficienti di trasmissione paterno e materno, che si leggono rispettivamente nell'ordinata e nell'ascissa del grafico della fig. 2, per i caratteri connessi alla religione non è distante dal 100%. Meno importante, ma sempre elevata, la determinazione verticale del comportamento politico: qui di solito padre e madre hanno ambedue una certa influenza. Molti altri comportamenti, in rapporto ad abitudini, convinzioni, superstizioni, preferenze, modi di divertirsi, sport, ecc., mostrano effetti più modesti della trasmissione dai genitori.
Uno studio simile compiuto su studenti di Taiwan ha dimostrato un'influenza anche maggiore della trasmissione verticale, ma naturalmente è difficile paragonare società così diverse tra loro come quelle americana e cinese. Estendendo, nei dati di Taiwan, l'analisi all'effetto di eventuali fratelli o sorelle maggiori si trova un'influenza paragonabile, e forse superiore, a quella dei genitori. In una società ancora molto diversa, quella dei Pigmei africani, in cui il gruppo sociale è numericamente ristretto (da 20 a 100 individui, raramente di più), non esistono scuole nè gerarchie sociali, le famiglie sono piccole e l'acculturazione termina in un'età abbastanza precoce, si trova che più dell'80% delle trasmissioni sono tra genitori e figli. Solo pochi tratti che di solito hanno un particolare significato sociale, come i costumi nel dividere la selvaggina, o le danze (sempre collettive e di notevole importanza sociale), vengono appresi largamente a livello del gruppo sociale. Pochissime innovazioni si sono diffuse per contagio.
È pure interessante considerare come alcuni caratteri o entità culturali tendono a mantenersi estremamente costanti, o comunque a mostrare poche variazioni nel tempo. La massima conservazione, abbiamo visto, è attesa quando la trasmissione è verticale, e ancor più chiaramente se è del tipo molti-a-uno, che tende a introdurre anche un'elevata uniformità nel comportamento individuale. In uno studio su oltre duecento società africane, di cui erano stati elencati oltre cinquanta caratteri nell'atlante etnografico di Murdock (v., 1967), Guglielmino-Matessi e altri (v., 1983) hanno potuto dimostrare che i caratteri più fortemente conservati sono quelli che si riferiscono alle regole di struttura della famiglia e della parentela. La dimostrazione è stata ottenuta in base a correlazioni dei caratteri culturali con la posizione nell'albero filogenetico linguistico. Anche se la lingua può talora essere bruscamente sostituita da un'altra, essa è pur sempre un ottimo indicatore dell'origine genetica di una popolazione e mostra di solito un'elevata correlazione con la posizione in un albero filogenetico (cladistico) costruito in base a dati genetici. Pur se mancano dati riguardanti direttamente questo specifico problema, è ragionevole accettare l'ipotesi che le strutture della famiglia e della parentela siano tipicamente trasmesse dai genitori e dagli altri membri del gruppo famigliare. Vi è anche un'elevata pressione del gruppo sociale e si realizzano quindi sia la trasmissione verticale che quella molti-a-uno, cioè i due tipi di trasmissione maggiormente responsabili di una elevata conservazione. Un altro esempio di notevole conservazione di una struttura famigliare è stato mostrato da recenti ricerche sulla famiglia nucleare inglese. La consuetudine in questo tipo di famiglia è che i figli lasciano presto la famiglia di origine, con la quale mantengono in seguito solo scarsi legami; si sposano e formano una nuova famiglia solo quando hanno raggiunto l'indipendenza economica. Demografi inglesi ritengono che questa struttura famigliare abbia stimolato il controllo delle nascite, in quanto la nuova famiglia tende a non avere figli se non può mantenerli; ha anche facilitato la genesi della società industriale, in quanto consente una maggiore mobilità geografica al singolo individuo, e quindi una maggiore efficienza nel seguire le opportunità di lavoro. Si asserisce addirittura che questa struttura famigliare sia anche responsabile dell'elevato individualismo caratteristico della società inglese. Ricerche storiche recenti hanno rintracciato l'esistenza della famiglia nucleare di tipo inglese nel XII secolo, e secondo demografi francesi essa sarebbe rintracciabile anche in gruppi etnici della Francia nordorientale (v. Todd, 1983).
Molti fatti demografici, e anche alcuni comportamenti politici rilevabili in base ai risultati delle elezioni politiche, sarebbero altamente correlati con le strutture famigliari tradizionali caratteristiche dei vari gruppi etnici. Tutte queste osservazioni fanno ritenere che le strutture famigliari siano particolarmente stabili sul piano evolutivo e che a ciò contribuiscano in modo importante i meccanismi della trasmissione culturale.
Val la pena infine di menzionare le ricerche di C. Marchetti che ha applicato un modello di selezione naturale a diversi esempi di selezione culturale. L'aumento della frequenza di un carattere biologico favorito dalla selezione naturale avviene secondo curve di tipo logistico (come la (1), che peraltro è stata introdotta in questo articolo per un processo diverso: la diffusione di tipo epidemico). È ben noto che curve come quelle generate dal processo definito dalla (1) si rettificano trasformando la proporzione di individui portatori del carattere, p, secondo la formula:
Y = log p/(1 − p).
L'espressione Y è quindi una funzione lineare del tempo.
Se la selezione è a favore di un carattere, si osserva una retta ascendente, con una pendenza che esprime il coefficiente di selezione; se a sfavore, la retta è discendente. Marchetti ha mostrato che le sorgenti di energia usate dall'uomo (v. fig. 3) sono soggette a una selezione che si comporta, almeno formalmente, come quella naturale. Così una nuova sorgente di energia - per esempio il carbone - ha soppiantato quella usata precedentemente, il legno, allo stesso modo che un mutante ne soppianta un altro nell'evoluzione biologica. È un esempio molto elegante di come la selezione culturale (cioè l'insieme delle nostre scelte e decisioni) può determinare la sostituzione graduale di un particolare prodotto o processo con un altro, così come la natura opera la sostituzione di un genotipo con un altro nel processo di selezione naturale. A sua volta il carbone dopo una fase ascendente è stato sostituito da un'altra sorgente di energia, il petrolio, ed è quindi entrato in una fase discendente. Le pendenze delle varie rette ascendenti e discendenti (v. fig. 3), essendo abbastanza simili, permettono di prevedere quando l'energia nucleare potrà soppiantare il petrolio (se ciò effettivamente avverrà), l'energia da fusione quella da fissione, ecc. Molti altri fenomeni culturali di tipo tecnologico sembrano seguire andamenti simili.
8. Interazioni fra genetica e cultura.
È difficile provare ma è anche di solito difficile escludere completamente la presenza della variazione genetica nella predisposizione individuale ad acquisire certi comportamenti, e perciò anche ad assorbire certi costumi, credenze, abitudini, che sono trasmessi culturalmente. L'interazione tra genetica e cultura che viene a crearsi è particolarmente complessa e difficile da analizzare, dato che abbiamo pochi mezzi per distinguere fra trasmissione genetica e culturale. La difficoltà è massima quando quest'ultima è prevalentemente del tipo verticale, che, come sappiamo, può simulare abbastanza accuratamente la trasmissione biologica.
Sono state compiute ricerche teoriche sulla competizione che può venire a crearsi tra la trasmissione biologica e quella genetica quando ambedue possono determinare lo stesso fenotipo. Si è visto che alla lunga la trasmissione biologica si impone, perché, a differenza di quella culturale, non commette errori, nel senso che, salvo mutazioni, essa permette una trasmissione perfetta, mentre è poco probabile che in una trasmissione culturale, diciamo verticale per fare un esempio, esattamente il 100% dei figli di genitori con un carattere porti anch'esso quel carattere. Lumsden e Wilson (v., 1981) hanno sostenuto una conclusione diversa, e cioè che la presenza della trasmissione culturale aiuta la fissazione di geni che permettono di ottenere lo stesso fenotipo realizzato per trasmissione culturale, ma il modello che essi hanno usato per raggiungere questa conclusione non ha finora avuto consensi.
La comparsa di una mutazione genetica che produce un fenotipo simile a quello realizzabile e trasmissibile per via culturale, e dotato di vantaggio selettivo, sarà quindi solitamente seguita dalla sua fissazione (anche se dopo un tempo molto lungo), ma non è peraltro necessario che in tal caso scompaia l'adattamento culturale preesistente. Possiamo allora capire come mai nella maggior parte degli animali superiori vi siano di solito sia l'insegnamento, spesso ma non esclusivamente materno, sia la predisposizione innata a sviluppare almeno alcune delle attività più necessarie alla sopravvivenza: la ricerca del cibo e la difesa dai pericoli. Talora la predisposizione innata è così sviluppata che è sufficiente a garantire un'elevata capacità di sopravvivenza anche se non c'è l'insegnamento materno. Ma sembra molto verosimile che vi sia vantaggio selettivo quando coesistono i due adattamenti, culturale e genetico.
Come abbiamo già detto varie volte, la più grande differenza tra la specie umana e tutte le altre sta nello sviluppo dei meccanismi culturali. È facile comprendere che meccanismi neuropsichici capaci di consentire un maggior adattamento culturale debbano aver avuto un notevole vantaggio selettivo, anche perché la possibilità di adattamento culturale consente la massima flessibilità nei confronti di ambienti e problemi assai diversi: può aiutare nell'adattamento sia a climi caldi che a climi freddi, nella caccia, nella pesca e in tutti i problemi pratici che vengono continuamente posti a ogni organismo vivente, sia a livello di sopravvivenza che di nutrizione e riproduzione. Inoltre questi problemi cambiano sovente nel tempo, donde il vantaggio di un sistema di adattamento particolarmente flessibile. Da una parte lo sviluppo particolare di meccanismi conoscitivi più avanzati può aver dato alla specie umana il vantaggio di una maggior capacità innovativa (abbiamo visto peraltro che anche in cugini non troppo vicini a noi la capacità di innovare non è del tutto assente); dall'altra, lo sviluppo del linguaggio, che nella specie umana è assai più avanzato che nelle altre, ha consentito di migliorare la velocità di trasmissione. Abbiamo visto che la fase della comunicazione è essenziale nel processo della trasmissione culturale, e chiaramente l'uomo, che ha la possibilità innata di imparare un linguaggio vasto e preciso, si trova nelle migliori condizioni per utilizzare in pieno i meccanismi di adattamento culturale e trasmetterli ai propri simili. Ciascuno di noi nasce con la capacità potenziale di apprendere in poco tempo conoscenze accumulate in millenni di storia culturale. Oggi il patrimonio culturale è ormai vastissimo, mentre i nostri meccanismi di apprendimento non si sono sviluppati in parallelo, così che ciascuno di noi acquisisce nel corso della vita una ben piccola parte del corredo attuale di conoscenze. La risposta alle complicazioni della vita culturale moderna è un'elevata specializzazione individuale, che naturalmente richiede scambi sociali sempre più complessi. I nostri sistemi di selezione culturale sono però rimasti immutati, ed è forse per questo motivo che sono divenuti inadeguati a rispondere in modo efficiente e tempestivo, e senza eccessive oscillazioni, alle nuove necessità che sorgono ogni momento.
Bibliografia.
Bonner, J. T., The evolution of culture in animals, Princeton 1980.
Boyd, R., Richerson, P. J., Culture and evolutionary process, Chicago 1985.
Campbell, D. T., On the conflicts between biological and social evolution and between psychology and moral tradition, in ‟American psychologist", 1975, XXX, pp. 1103-1126.
Cavalli-Sforza, L. L., Feldman, M. W., Cultural transmission and evolution: a quantitative approach, Princeton 1981.
Cavalli-Sforza, L. L., Feldman, M. W., Cultural versus genetic adaptation, in ‟Proceedings of the National Academy of Science", 1983, LXXX, pp. 4993-4996.
Cavalli-Sforza, L. L., Feldman, M. W., Chen, K. H., Dornbusch, S. M., Theory and observation in cultural transmission, in ‟Science", 1982, CCXVIII, pp. 19-27.
Feldman, M. W., Cavalli-Sforza, L. L., Cultural and biological evolutionary process: gene-culture disequilibrium, in ‟Proceedings of the National Academy of Science", 1984, LXXXI, pp. 1604-1607.
Guglielmino-Matessi, R. C., Viganotti, C., Cavalli-Sforza, L. L., Spatial distributions and correlations of cultural traits in Africa, Istituto di Analisi Numerica del Consiglio Nazionale delle Ricerche, Pavia 1983.
Hagerstrand, T., Innovation diffusion as a spatial process, Chicago-London 1967.
Kroeber, A. L., Kluckhorn, C., Culture, a critical review of the concepts and definitions, in ‟Papers of the Peabody Museum of American Archaeology and Ethnography", 1952, XLVII, 1, pp. 1-233.
Lorenz, K., Der Kumpan in der Umwelt des Vogels, in ‟Journal of ornithology", 1935, LXXXIII, pp. 137-413.
Lumsden, C., Wilson, E. O., Genes, mind and culture, Cambridge, Mass., 1981.
Mainardi, D., L'animale culturale, Milano 1974.
Mainardi, D., Tradition and social transmission of behavior in animals, in Sociobiology: beyond nature-nurture? (a cura di G. W. Barlow e J. Silverberg), Boulder, Colo., 1980.
Marchetti, C., Society as a learning system: discovery, invention, and innovation cycles revisited, in ‟Technological forecasting and social change", 1980, XVIII, pp. 267-282.
Marchetti, C., Nakicenovic, N., The dynamics of energy systems and the logistic substitution model, International Institute for Applied Systems Analysis - IIASA, Laxenburg, Austria, RR-79-13, December 1979.
Murdock, G. P., Ethnographic atlas, Pittsburgh 1967.
Rogers, E. M., Shoemaker, F. F., Communication of innovations: a cross-cultural approach, New York 1971.
Todd, E., La troisième planète, Paris 1983 (tr. it.: Il terzo pianeta, Roma 1983).
White, L. A., The evolution of culture, New York 1959.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


