Evoluzione genetica dell'uomo
Enciclopedia della Scienza e della Tecnica (2007)
Evoluzione genetica dell'uomo
Che cosa è l'uomo, se il suo supremo bene e tutto il prezzo del suo tempo restringesi nel mangiare e nel dormire? Un bruto, e null'altro. Certo quegli che ne dotò di questa sublime ragione, che può veder nel passato e nell'avvenire, non ci ha data questa intelligenza, celeste facoltà, perch'ella in noi si rimanga inerte. Questo si legge nell'Amleto di William Shakespeare.
Comprendere la base genetica dei tratti fisici e comportamentali che distinguono gli esseri umani dagli altri Primati rappresenta una delle nuove grandi sfide della biologia. Dei milioni di differenze nelle coppie di basi tra esseri umani e scimpanzé, quali particolari cambiamenti hanno contribuito all'evoluzione delle caratteristiche dell'uomo in seguito alla separazione delle discendenze Pan e Homo qualche milione di anni fa? Cosa rende gli esseri umani moderni diversi dalle grandi scimmie antropomorfe e dai primi Ominidi? In quali Ominidi e quando nel processo evolutivo sono apparsi i tratti fisici e i comportamenti significativi? Dove, nei nostri cervelli più grandi, risiedono le facoltà specifiche degli esseri umani? Si tratta di interrogativi di vecchia data in paleoantropologia e in anatomia comparata, fin dai tempi della scoperta dei teschi di Neanderthal e dai primi studi sulle grandi scimmie antropomorfe nel XIX secolo. Oggi risolvere il mistero delle origini dell'uomo significa andare oltre la descrizione e la storia dei suoi tratti distintivi, indagando i meccanismi genetici che stanno alla base della loro formazione ed evoluzione. Con la caratterizzazione del genoma umano, e quella del nostro 'cugino' scimpanzé in arrivo, le indagini per scoprire la base genetica dei tratti fisici e comportamentali che ci distinguono dalle altre scimmie antropomorfe stanno rapidamente acquistando slancio.
I genomi divergono in funzione del tempo e la maggior parte dei cambiamenti che si accumulano nella sequenza tra due qualsiasi specie imparentate tra loro sono selettivamente neutri o quasi neutri, poiché non sembrano contribuire a creare differenze funzionali o fenotipiche che possano costituire dei vantaggi adattativi. È molto importante sviluppare metodi per distinguere le 'prove incontrovertibili' dell'evoluzione genetica dell'uomo, dovuta soprattutto alla selezione naturale, dal normale e non significativo ticchettio dell'orologio dell'evoluzione molecolare. La misura e la velocità dell'evoluzione morfologica negli Ominidi suggeriscono molti cambiamenti evolutivi indipendenti e incrementali che, sulla base delle recenti scoperte in animali modello, si ritiene siano di natura poligenica e regolatrice. La genomica comparativa, la genetica delle popolazioni, le analisi dell'espressione genica e la genetica medica hanno iniziato a fare incursioni, in modo sinergico, nella complessa architettura dell'evoluzione umana. La grande sfida consiste nello spiegare il numero, l'identità e le funzioni dei geni, nonché i cambiamenti specifici al loro interno, che hanno dato forma all'evoluzione dei tratti distintivi del genere Homo e in particolare di Homo sapiens. Questo traguardo è stato realizzato solo per alcuni tratti distintivi in sistemi modello: si tratta infatti di un compito difficile applicare questo tipo di indagine alle caratteristiche umane sulle quali sappiamo così poco, e in una prospettiva ampia quanto l'intero arco dell'evoluzione umana.
L'albero degli Ominidi
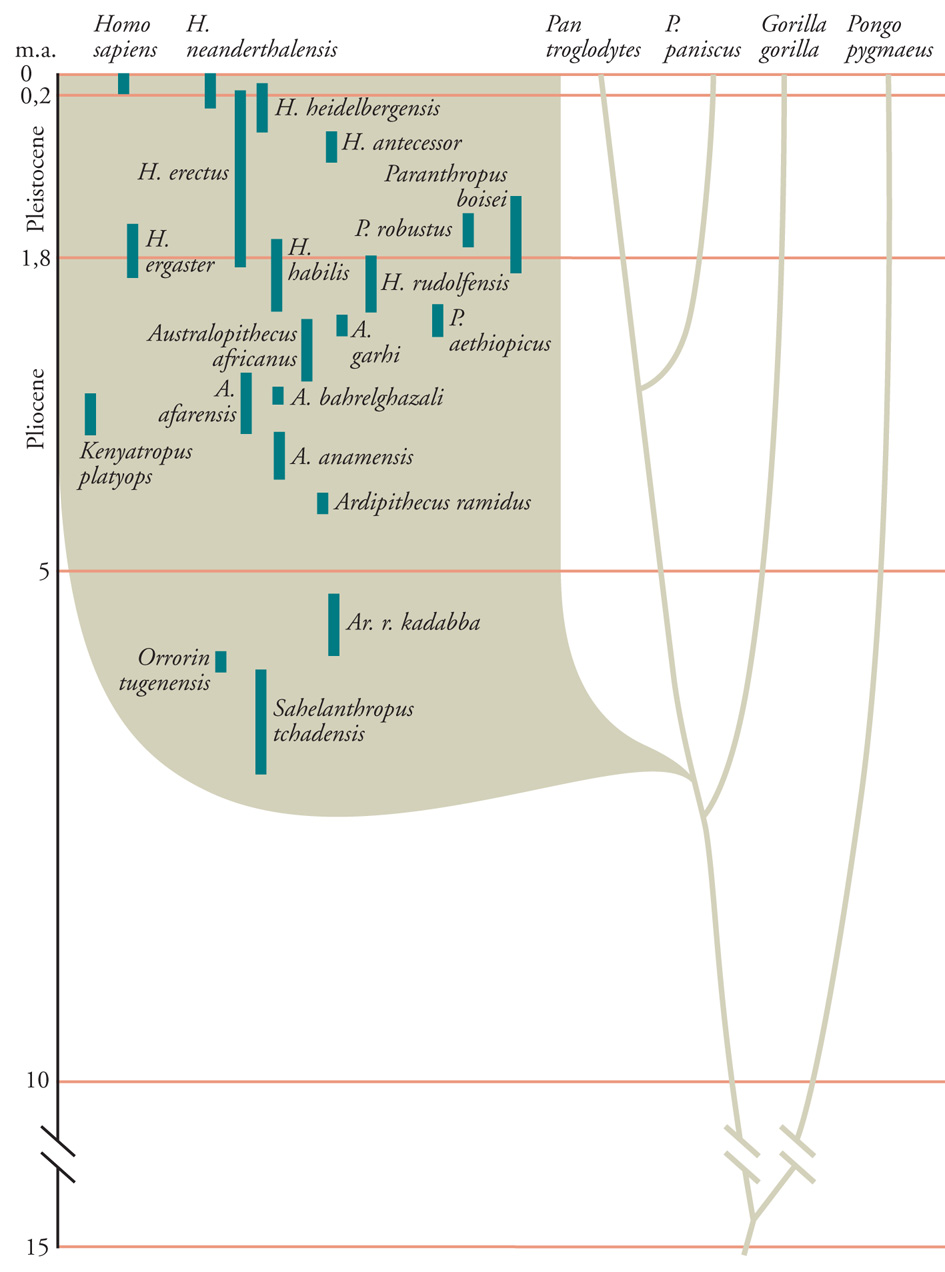
Per avvicinarsi alle origini dei tratti distintivi dell'uomo a livello genetico, è essenziale avere come sistema di riferimento una storia della nostra discendenza e dei caratteri che la distinguono. È inadeguato e fuorviante considerare soltanto l'anatomia e lo sviluppo comparativi (o i genomi) degli esseri umani, degli scimpanzé e delle altre scimmie antropomorfe ancora esistenti, e poi tentare di dedurre come le differenze esistenti potrebbero essere codificate ed espresse. Ciascuna di queste specie ha una discendenza indipendente che risale a quella degli Ominini o persino più indietro nel tempo, il termine Ominini si riferisce agli esseri umani e ai nostri antenati nella scala dell'evoluzione, risalenti al tempo della separazione della discendenza umana da quella delle scimmie antropomorfe; Ominidi è riferito agli esseri umani e alle scimmie antropomorfe africane (fig. 2). L'evoluzione dei tratti distintivi moderni non fu un processo cumulativo, lineare, e le idee sul ritmo, sul modello e sulla grandezza del cambiamento possono essere verificate solo attraverso i reperti fossili, che però sono sempre suscettibili di revisione in seguito a nuovi ritrovamenti. Le testimonianze fossili continuano a determinare i punti di vista su tre questioni cruciali relative all'evoluzione degli Ominidi. Primo, cosa distingue gli Ominini dalle scimmie antropomorfe? Secondo, cosa distingue gli esseri umani moderni (Homo sapiens) dai primi Ominini? E terzo, quale era la natura dell'ultimo antenato comune degli Ominini e della discendenza Pan?
Le conclusioni circa l'ordine cronologico e la grandezza dell'evoluzione dei caratteri degli Ominini dipendono da un modello di albero della loro evoluzione. Si tratta di una questione controversa e tuttora irrisolta, in parte a causa della frequenza degli entusiasmanti ritrovamenti fossili nel corso degli ultimi due decenni. Un punto di vista che si sta facendo strada rappresenta l'evoluzione degli Ominini come una serie di radiazioni adattative durante le quali ebbero origine molte diverse ramificazioni della discendenza degli Ominini, che però si estinsero (fig. 2). Uno dei presupposti di questo modello è che varie caratteristiche anatomiche si sarebbero ritrovate in diverse combinazioni negli Ominini per effetto della loro indipendente acquisizione, modificazione e perdita in specie diverse. Per esempio, la scoperta di un cranio fossile di Sahelanthropus tchadensis, risalente a 6-7 milioni di anni fa, che presentava un cervello delle dimensioni di uno scimpanzé ma caratteristiche facciali e dentali simili a quelle di un ominine, rappresenta il tipo di morfologia che sarebbe compatibile con una radiazione di animali simili a scimmie antropomorfe, da cui affioravano le linee principali della discendenza ominina (anche se l'interpretazione delle affinità di questo fossile è controversa).
Tendenze evolutive negli Ominini fossili
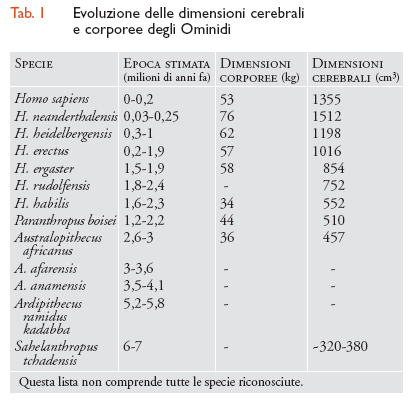
In teoria, se si identificassero tutte le specie umane e di scimmie antropomorfe fossili e se avessimo a disposizione molti esemplari quasi completi si potrebbe ricostruire la comparsa degli aspetti distintivi dell'uomo e dello scimpanzé nel corso del tempo. Ma per la maggior parte delle discendenze ciò non accade; infatti non sono stati identificati fossili di scimpanzé arcaici. Dobbiamo accontentarci di un quadro parziale e spesso confuso dell'evoluzione dei tratti distintivi dell'uomo. Tra tutta una serie di caratteristiche qualitative e quantitative che distinguono gli esseri umani da altre scimmie antropomorfe (tab. 1), il cervello grande, il bipedismo, i canini piccoli, le avanzate capacità di utilizzare il linguaggio e di realizzare degli utensili sono stati al centro dell'attenzione della paleoantropologia. I tratti fisici principali non sono generalmente degli elementi singoli, ma richiedono dei cambiamenti concomitanti negli elementi scheletrici coinvolti nella locomozione (per es., nella colonna vertebrale, nel bacino e nei piedi, e nelle proporzioni degli arti), nella presa (la morfologia della mano e un pollice allungato e opponibile) e nella masticazione del cibo (la mandibola e la dentizione), come anche i tratti relativi al ciclo vitale come la lunghezza della vita. È una fortuna che la maggior parte delle caratteristiche scheletriche si prestino a dettagliati studi quantitativi delle testimonianze fossili.
Tra gli Ominini, alcune tendenze nell'evoluzione della grandezza del corpo, del cervello e della dentizione sono evidenti. Le specie più recenti sono caratterizzate da una massa corporea maggiore, dei cervelli relativamente più grandi, gambe più lunghe rispetto al tronco e denti più piccoli, mentre le specie più antiche avevano in generale cervelli e corpi di dimensioni inferiori (tab. 1), gambe più corte rispetto al tronco e denti più grandi. Questi tratti caratteristici vanno evidenziati per concentrare l'attenzione sulla grandezza e sulla cronologia dell'evoluzione dei caratteri, e sul numero (crescente) di taxa complessivamente riconosciuti relativi agli Ominini. Qualunque sia stato lo schema di ramificazione dell'albero degli Ominini, i cambiamenti relativi sostanziali hanno avuto luogo in un arco di tempo prolungato e nel corso di un significativo numero di eventi di speciazione. Ci fu un marcato aumento della dimensione assoluta del cervello a partire dal Pleistocene inferiore e di nuovo nel Pleistocene medio, con un lungo intervallo di forse un milione di anni durante il quale le dimensioni del cervello non cambiarono in modo significativo. Per quanto riguarda Homo sapiens moderno, è interessante notare che in Homo neanderthalensis le dimensioni del corpo e del cervello erano persino maggiori; non esiste alcuna ovvia spiegazione fisica che giustifichi il successo di Homo sapiens e la scomparsa dell'Homo neanderthalensis.
Una mente meravigliosa
L'incremento relativo nelle dimensioni del cervello, seppure marcato, rappresenta soltanto un'indicazione generica di un potenziale aumento delle capacità cognitive. Dal momento che ci si è resi conto già da molto tempo che esistono aree distinte del cervello preposte all'elaborazione delle diverse funzioni cognitive, motorie e sensoriali, i neuroanatomisti comparativi hanno tentato di identificare quelle aree che potrebbero essere centrali per l'evoluzione delle capacità umane. C'è una vecchia teoria secondo la quale la corteccia frontale (coinvolta nella pianificazione, nell'organizzazione, nella personalità, nel comportamento e in altre funzioni cognitive 'più alte') sarebbe sproporzionatamente più grande negli esseri umani, ma sembra che non sia così (è più grande, ma non in modo eccessivo). Poiché le evidenti differenze anatomiche non spiegano le capacità cognitive, si è andati alla ricerca delle differenze relative nelle dimensioni, nella composizione cellulare, nella dettagliata citoarchitettura e/o connettività delle aree del cervello degli esseri umani e delle grandi scimmie antropomorfe, per spiegare la comparsa delle capacità umane.
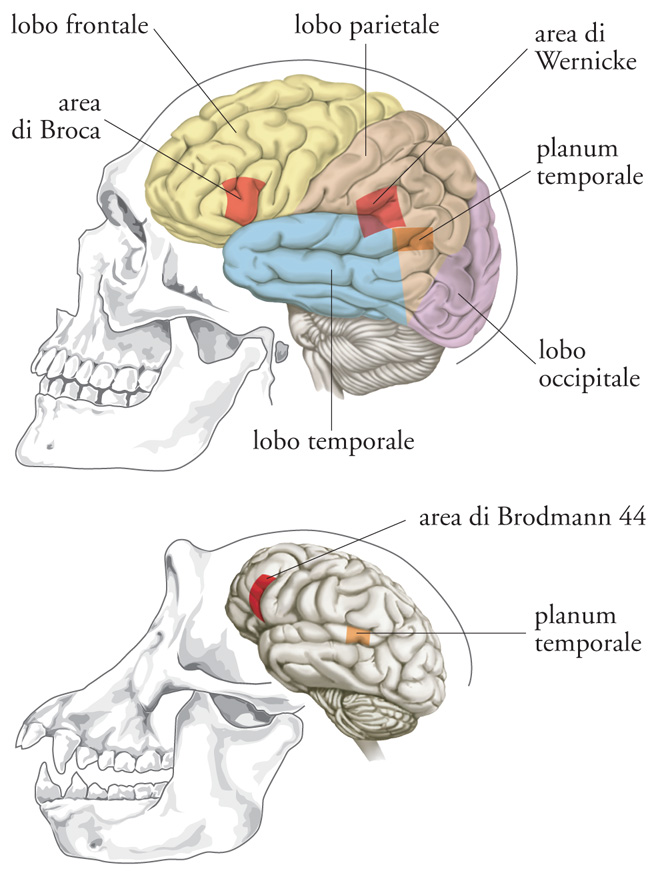
Di sommo interesse sono la produzione e la comprensione del linguaggio. Due aree in particolare hanno suscitato l'attenzione maggiore. Una è l'area di Broca nel lobo frontale della neocorteccia (fig. 3). Questa regione è più ampia nell'emisfero sinistro del cervello rispetto a quello destro, una asimmetria che è stata correlata con la facoltà del linguaggio. Grazie alle immagini alla risonanza magnetica di scimpanzé, bonobo e gorilla, in queste grandi scimmie antropomorfe è stata individuata una asimmetria sinistra-destra simile. Ciò indica che il substrato neuroanatomico della dominanza dell'emisfero sinistro nella elaborazione del linguaggio precedette la nascita degli Ominini. L'emisfero sinistro, inoltre, controlla di solito il destrismo: è perciò interessante notare come nelle scimmie antropomorfe in cattività i gesti manuali siano influenzati dalla mano destra e come questa tendenza risulti aumentata quando la vocalizzazione si combina all'espressione gestuale, a indicare un processo di comunicazione controllato dall'emisfero sinistro.
Una seconda area di interesse è la ricettiva area di Wernicke, posizionata posteriormente nel lobo temporale e preposta al linguaggio (fig. 3). Un luogo all'interno di quest'area, il planum temporale, è coinvolto nella comunicazione umana (sia parlata sia gestuale) e nell'attitudine musicale, e anche questo mostra una dominanza dell'emisfero sinistro. Nella maggior parte degli esseri umani, la scissura di Silvio associata al planum temporale sinistro si estende più posteriormente. Prova di questa asimmetria si è rintracciata nei calchi endocranici fossili di H. habilis, H. erectus e H. neanderthalensis. Cosa ancora più importante, l'esistenza di una tipologia asimmetrica di planum temporale è stata di recente individuata negli scimpanzé (fig. 3).
Sono state avanzate parecchie ipotesi per spiegare la presenza nelle grandi scimmie antropomorfe di queste due caratteristiche neuroanatomiche associate alla comunicazione umana. Sebbene sia possibile che siano comparse e abbiano acquisito le proprie funzioni in modo indipendente in ciascuna discendenza, la spiegazione più semplice è che l'antenato comune delle grandi scimmie antropomorfe e degli esseri umani avesse dei centri asimmetrici che erano coinvolti nella comunicazione, e che queste strutture abbiano subito delle modificazioni evolutive indipendenti negli scimpanzé e negli Ominini. Se fosse davvero così, allora la sfida a cui la neuroanatomia comparativa si troverebbe di fronte consisterebbe nell'identificare ulteriori sottili differenze nella sottorganizzazione (cioè nella microanatomia) che interessano le interconnessioni delle regioni corticali, nella parte circuitale locale e/o nella citoarchitettura, e che potrebbero essere presenti solo nei cervelli umani. Recentemente, si è scoperto che le dimensioni delle colonne verticali di neuroni presenti nella corteccia, noti come 'minicolonne', differiscono tra esseri umani e scimpanzé nel planum temporale. Inoltre, è stato dimostrato che l'area 10 della corteccia prefrontale, che è coinvolta nelle funzioni cognitive più alte, è più estesa e specializzata negli esseri umani rispetto alle scimmie antropomorfe. Queste osservazioni suggeriscono che le capacità umane siano più il prodotto dei cambiamenti quantitativi nelle aree specializzate che delle novità neuroanatomiche.
Sviluppo delle caratteristiche degli Ominidi
Le differenze morfologiche tra gli esseri umani moderni, i primi Ominini e le grandi scimmie antropomorfe sono, naturalmente, il prodotto di cambiamenti intercorsi durante lo sviluppo. Studi comparativi sullo sviluppo dei crani umani e di scimpanzé, esami straordinariamente dettagliati sull'ontogenesi craniofacciale di H. neanderthalensis e sulla formazione della dentatura negli antichi Ominini, hanno consentito di comprendere un gran numero di cambiamenti evolutivi che sono alla base della dimensione e della morfologia dei crani degli esseri umani moderni.
Una delle differenze fondamentali da lungo tempo apprezzate nello sviluppo degli scimpanzé e degli esseri umani è la velocità relativa di crescita e maturazione del cranio. I neonati umani hanno crani meno maturi dal punto di vista della forma rispetto ai giovani scimpanzé, ma crani (e cervelli) molto più grandi. Questi vengono tradizionalmente descritti come cambiamenti eterocronici, i quali generano caratteri neotenici nei quali la maturazione viene ritardata, le dimensioni aumentano e la forma assomiglia alle forme giovanili degli antenati. Alla fine i crani degli scimpanzé e degli esseri umani arrivano ad avere le stesse dimensioni, mostrando ulteriori modificazioni relative ai periodi di crescita infantile e adolescenziale e marcate differenze nelle dimensioni del volto e nel volume cerebrale. È importante notare che tutti i mutamenti scheletrici associati al bipedismo sono innovazioni strutturali indipendenti dalla neotenia. Queste osservazioni suggeriscono che il cervello umano non è il prodotto di semplici cambiamenti nelle relazioni di crescita, ma di molteplici modifiche indipendenti e sovrapposte.
Rispetto a specie di Ominini più recenti, la forma craniofacciale degli esseri umani moderni sembra essere stata modellata dai cambiamenti negli elementi che influenzano la posizione spaziale del volto, del neurocranio e della base cranica. Gli esseri umani moderni sono caratterizzati da una volta cranica più arrotondata e da retrazione facciale (la posizione anteroposteriore del volto in rapporto alla base del cranio e al neurocranio). Raffronti con lo sviluppo cranico di H. neanderthalensis desunti da fossili di individui di età differente, fanno pensare che le tipiche differenze craniche tra le caratteristiche di H. neanderthalensis e dell'essere umano moderno compaiano in una fase iniziale dell'ontogenesi.
L'infanzia prolungata, la ritardata maturazione sessuale e la lunghezza della nostra vita sono tratti del ciclo vitale che determinano aspetti importanti della società umana. Spunti per comprendere lo sviluppo di questi cambiamenti evolutivi nel corso dell'evoluzione vengono offerti da una dettagliata analisi comparativa dello sviluppo dei denti, il quale è correlato agli stadi di crescita e di sviluppo dei Primati. I ritmi di formazione dello smalto in Ominini fossili suggeriscono che i tempi di sviluppo dentale fossero più brevi negli Australopitechi e nei primi membri del genere Homo rispetto agli esseri umani moderni. Ciò indica che lo schema moderno di sviluppo dentale e i tratti evolutivi a esso associati apparvero in una fase tarda dell'evoluzione umana. Quando lo consideriamo nel contesto degli altri tratti distintivi, come le dimensioni cerebrali e le proporzioni corporee, emerge un modello di evoluzione a mosaico, con tratti differenti che compaiono in momenti diversi e forse in combinazioni diverse nella storia degli Ominini.
L'evoluzione umana rappresentò un'eccezione?
L'importanza, la velocità e lo schema del cambiamento durante l'evoluzione degli Ominini, dedotti dalle testimonianze fossili, dalla neuroanatomia e dall'embriologia comparative, forniscono il fondamento essenziale per avvicinarsi alla genetica dell'evoluzione umana. Dagli studi sopra discussi emergono cinque punti chiave che sono in relazione con i tentativi di ricostruire gli eventi genetici che stanno alla base dell'origine e delle modificazioni dei tratti caratteristici degli esseri umani. Per prima cosa, l'evoluzione dei tratti caratteristici non fu lineare. L'incremento di ~1000 cm3 nelle dimensioni cerebrali nel corso di 5-7 milioni di anni non avvenne alla stessa velocità relativa nella filogenesi degli Ominini: a volte fu molto lento, e poi più veloce in alcuni intervalli, e in periodi più recenti sembra tornato a uno stadio precedente. In secondo luogo, la maggior parte dell'evoluzione dei tratti caratteristici consiste in semplici cambiamenti quantitativi (cioè, i tratti caratteristici sono continui). In terzo luogo, i ritmi evolutivi non furono affatto eccezionali in merito all'evoluzione dei Mammiferi. Ad esempio, le discendenze equine fossili del tardo Pliocene-Pleistocene mostrano ritmi di cambiamento nella dimensione corporea e nelle altre caratteristiche simili a quelle degli Ominidi. In quarto luogo, buona parte dei cambiamenti evolutivi precedettero l'origine del genere Homo e di H. sapiens: la storia della nostra specie rappresenta appena l'ultimo 3% dell'intervallo temporale dell'evoluzione degli Ominini. In quinto luogo, infine, molte caratteristiche sono presenti non solo negli esseri umani ma anche nelle scimmie antropomorfe. Ciò vuol dire che alla base della maggior parte dell'evoluzione umana c'è la modifica delle strutture esistenti e dei percorsi evolutivi, piuttosto che l'invenzione di nuove caratteristiche. Queste osservazioni indicano che l'evoluzione morfologica negli Ominini non rappresentò un'eccezione, ma fu il prodotto di cambiamenti genetici ed evolutivi tipici di altri Mammiferi e animali.
Architettura genetica dell'evoluzione dei tratti distintivi
Date le dimensioni dell'evoluzione degli Ominini, dedotta dalle testimonianze fossili e dall'anatomia comparata, cosa possiamo aspettarci per ciò che riguarda la complessità genetica che sta alla base dell'evoluzione dei tratti distintivi? Per esempio, secondo una vecchia e consolidata tendenza, quegli eventi che in base alla documentazione fossile sono percepiti come relativamente rapidi vengono ascritti a una o forse a più mutazioni radicali, e tra questi c'è anche la recente evoluzione dell'uomo. L'incremento relativo delle dimensioni cerebrali nell'arco di 5 milioni di anni, o la sua estesa funzione cognitiva, potrebbero essere dovuti soltanto a uno o ad alcuni cambiamenti genetici? I migliori criteri guida (e, al momento, gli unici a disposizione) per rispondere a questo quesito provengono da dettagliati studi di genetica su organismi modello, che sono riusciti a esaminare minuziosamente la genetica della formazione, variazione ed evoluzione dei tratti complessi.
Nei sistemi modello si sono stabiliti sei essenziali concetti generali che riguardano la potenziale architettura genetica dell'evoluzione dei tratti distintivi dell'uomo: (a) la variazione nei tratti quantitativi e continui è generalmente poligenica. Studi sulla variazione in specie modello rivelano che molti geni hanno scarso effetto, mentre a volte uno o più geni sono di grande effetto e controllano i parametri dei tratti. Negli esseri umani, uno studio della variazione in 20 variabili antropometriche condotto in due diversi gruppi etnici ha rivelato che più del 50% della variazione è poligenica; (b) la velocità di evoluzione dei tratti distintivi non ci dice nulla circa il numero di geni coinvolti. Studi sulla selezione artificiale e sulla divergenza interspecifica indicano che l'intensità della selezione e l'ereditabilità sono determinanti dalla velocità evolutiva più importanti di quanto non sia la complessità genetica dei tratti distintivi sottoposti a selezione. Esiste una considerevole variazione costante nei tratti distintivi, comprese caratteristiche che si potrebbero considerare altamente vincolate, come la morfologia degli arti nei Tetrapodi. In generale, i ritmi evolutivi che si sono osservati sotto l'effetto della selezione naturale sono molto più lenti di quanto non sia potenzialmente possibile. La variazione genetica o la complessità genetica non è il fattore limitante; anzi, la considerevole variazione genetica sta persino alla base dei tratti fenotipicamente invarianti. Dal momento che la velocità a cui un tratto distintivo appare nella documentazione fossile non ci dice nulla circa l'architettura genetica, si deve resistere alla tentazione di appellarsi a modelli macromutazionali per i cambiamenti rapidi in assenza di prove genetiche; (c) la variazione e la divergenza morfologiche sono associate ai geni che regolano lo sviluppo. Confronti tra le basi evolutive dello sviluppo dello schema corporeo negli animali suggeriscono che l'evoluzione morfologica è il prodotto di cambiamenti nella distribuzione spazio-temporale dei geni regolatori e dell'evoluzione delle reti regolatorie dei diversi geni. Si ritiene che i cambiamenti evolutivi nella discendenza umana siano associati a geni che influenzano i parametri evolutivi, come quelli che codificano i fattori di trascrizione e gli elementi dei percorsi di trasduzione del segnale; (d) le mutazioni responsabili della variazione dei tratti distintivi sono spesso in regioni regolatorie non codificanti. Quando è stato possibile identificare la variazione in geni che stanno alla base di variazioni fenotipiche o di differenze nei livelli proteici, si è visto che la causa della variazione è spesso ascrivibile a inserzioni o sostituzioni nelle regioni regolatorie e nelle regioni non codificanti; (e) le sostituzioni multiple del nucleotide spesso caratterizzano gli alleli. Analisi dettagliate dei loci per i tratti quantitativi hanno spesso mostrato che le differenze funzionali tra gli alleli sono dovute a differenze multiple del nucleotide; esse indicano inoltre che le interazioni non additive tra i siti all'interno di un locus potrebbero essere la chiave per il differenziamento degli alleli, e che il contributo di ogni singolo sito potrebbe essere modesto (e difficile da scoprire); (f) esiste una qualche concordanza tra i geni responsabili della variazione intraspecifica e della divergenza interspecifica. L'analisi genetica della divergenza interspecifica è possibile solamente in certe circostanze, quando la riproduzione in laboratorio riesce a superare gli ostacoli dovuti alla specie e si può fare una mappatura dei tratti distintivi. In alcuni casi, si è scoperto che alcuni degli stessi loci sono coinvolti sia nella variazione all'interno della specie sia nella divergenza tra specie. Ciò fa sperare che gli studi sulla variazione intraspecifica negli esseri umani possano portare alla scoperta di geni che sono stati importanti nella storia dell'umanità.
Dal momento che l'evoluzione dei tratti distintivi dell'uomo ha seguito un corso incrementale del tutto simile a quello dei tratti distintivi studiati in sistemi modello, questi sei concetti indicano che dovremmo aspettarci una base altamente poligenica per tratti complessi come le dimensioni cerebrali, la morfologia e lo sviluppo craniofacciale, le aree corticali della parola e del linguaggio, la morfologia della mano e delle dita, la morfologia dei denti e dello scheletro postcranico. Dovremmo inoltre anticipare che i molteplici cambiamenti nelle regioni regolatorie non codificanti e nei geni regolatori sono di grande importanza. Ma come facciamo a trovarli?
L'aritmetica dell'evoluzione della sequenza umana
Tutti gli approcci genetici alle origini dell'uomo sono fondamentalmente comparativi e cercano di identificare i cambiamenti genetici che hanno avuto luogo in particolare nella discendenza umana e che hanno contribuito alla differenziazione degli esseri umani dal nostro ultimo antenato comune con le scimmie antropomorfe o con altre specie di Homo. Il riferimento comparativo principale è il genoma dello scimpanzé (Pan troglodytes), il nostro parente più prossimo tra gli esseri viventi, con cui condividiamo un antenato che visse 5-7 milioni di anni fa. Una buona parte delle differenze esistenti tra i due genomi potrebbero essere costituite dagli elementi trascurabili delle sostituzioni neutre e dall'aumento e dalla perdita di elementi ripetitivi nel corso di lunghi periodi di tempo (più del 46% del DNA umano è composto da ripetizioni distribuite a intervalli), ma una piccola parte dei cambiamenti nella sequenza genomica è responsabile delle differenze ereditarie tra le specie. Il punto nodale della sfida sta nel come distinguere quei cambiamenti specifici che sono biologicamente significativi dai molti che non lo sono.
Nel caso dell'evoluzione umana, ci sono tre questioni genetiche fondamentali che vorremmo cercare di comprendere. Primo, quanti geni furono direttamente coinvolti nella nascita dell'anatomia, della fisiologia e del comportamento umani (alcuni, dozzine, centinaia o migliaia)? Secondo, quali geni specifici contribuirono alla nascita di particolari tratti distintivi dell'uomo? E terzo, che tipo di cambiamenti in questi geni contribuirono all'evoluzione (per es., duplicazioni dei geni, sostituzioni degli amminoacidi o evoluzione della sequenza di regolazione)? Nei pochi pionieristici studi che si stanno occupando direttamente della base genetica della divergenza tra uomo e scimpanzé, si sono perseguite strategie differenti ma in qualche modo complementari che stanno iniziando a mettere in luce la portata dell'evoluzione genetica dell'uomo e, in alcuni casi, geni specifici che potrebbero essere stati sottoposti a selezione nel corso della recente evoluzione umana.
Genomica comparativa
Le differenze tra i genomi animali rilevate più facilmente sono espansioni o contrazioni di famiglie di geni. Sebbene il genoma completo dello scimpanzé non sia ancora disponibile, una parziale mappa comparativa indica che ci sono regioni del genoma umano che potrebbero non essere rappresentate negli scimpanzé o in altre scimmie antropomorfe. Tali regioni potrebbero essere dovute a duplicazioni o inserzioni che ebbero luogo nella discendenza degli Ominini o a delezioni nella discendenza degli scimpanzé. Una famiglia di geni, denominata morpheus, subì un processo di espansione nel corso di una duplicazione segmentale sul cromosoma umano 16. Questa espansione è condivisa da altre grandi scimmie antropomorfe, ma sembra che ci fossero anche duplicazioni che sono specifiche della discendenza umana.
Sulla base dei confronti con altri genomi, in particolare con la sequenza del topo, tali duplicazioni specifiche delle discendenze non sono una sorpresa. Nei 75 milioni di anni o più intercorsi dalla separazione dell'antenato comune dei topi e degli esseri umani, si formarono parecchie dozzine di gruppi di geni, specifici del topo, che sono generalmente rappresentati da un singolo gene nel genoma umano. Il fatto che gli esseri umani e le scimmie antropomorfe si siano separati in tempi più recenti suggerisce che la serie genica specifica degli esseri umani sarà più esigua. È interessante notare che una parte significativa del gruppo genico del topo codifica proteine che giocano un ruolo nella riproduzione, nell'immunità e nell'olfatto. Ciò indica che la selezione sessuale, gli agenti patogeni e l'ecologia sono in grado di dare forma alle principali differenze tra i Mammiferi nel contenuto del codice genetico. Si deve inoltre notare che l'80% dei geni dei topi hanno un ortologo di 1:1 nel genoma umano, e che più del 99% hanno un qualche omologo. Queste cifre e questi dati sintetici suggeriscono che esiste un repertorio genico che è qualitativamente quasi identico tra i Mammiferi. La presenza o assenza di particolari duplicati genici potrebbe riflettere un cambiamento causato dall'adattamento, ma saranno necessarie ulteriori prove per determinare se sui geni abbia agito la selezione positiva.
Migliaia di cambiamenti adattativi nel proteoma umano?
Il primo luogo in cui si sono cercati i cambiamenti genetici adattativi è nelle sequenze codificanti per le proteine. Se le sostituzioni che hanno caratterizzato l'evoluzione umana sono uniformemente distribuite attraverso tutto il genoma, ci si aspetterà che solo una piccola parte cada all'interno delle regioni codificanti, parte delle quali potrebbe inoltre essere sostituzioni sinonimiche. Si sono sviluppati vari metodi per rilevare se le sostituzioni degli amminoacidi potessero essere il risultato della selezione positiva, cioè dell'evoluzione adattativa. Per stimare la misura della selezione positiva nell'evoluzione proteica umana, sono stati esaminati i dati relativi alla divergenza nella sequenza per 182 geni umani e delle scimmie del Vecchio Mondo, nonché i dati relativi al polimorfismo per un numero simile di geni umani. Prendendo in considerazione la frequenza dei polimorfismi comuni (ignorando gli alleli rari), è stato osservato un grado di sostituzioni degli amminoacidi più grande di quanto non ci si aspettasse, la qual cosa costituisce la prova della selezione.
La proiezione sull'intero proteoma ha stimato che il 35% delle sostituzioni degli amminoacidi tra l'uomo e le scimmie del Vecchio Mondo sono state causate dalla selezione positiva. Questa proporzione è sostanzialmente maggiore di quanto non ci si aspetterebbe se la maggior parte delle mutazioni fossero neutre o quasi neutre. Se si tratta dell'ordine di grandezza corretto, rappresenta un vero e proprio incubo per l'identificazione dei geni chiave sottoposti a selezione, perché questa cifra suggerisce che, in media, negli ultimi 5 milioni di anni in ogni proteina umana abbiano avuto luogo due o più sostituzioni adattative. È possibile che la cifra, basata sullo studio di meno dello 0,5% del proteoma umano, rappresenti una sovrastima della parte o della distribuzione delle sostituzioni adattive. È chiaro che alcune proteine siano sottoposte a una forte pressione per rimanere costanti, mentre altre, specialmente quelle coinvolte nelle cosiddette 'corse agli armamenti molecolari', sono sottoposte a pressione per cambiare. Per esempio, le proteine del complesso maggiore di istocompatibilità, che interagiscono con sostanze estranee varie e sempre diverse, mostrano chiari segni di selezione. Le proteine coinvolte nella riproduzione, che giocano un ruolo nella competizione spermatica o nel riconoscimento tra i gameti, sembrano anch'esse evolvere più velocemente e sotto l'azione di un certo grado di selezione positiva. Un gran numero di proteine umane della riproduzione maschile hanno un livello superiore alla media di sostituzioni degli amminoacidi. Sebbene l'evoluzione accelerata delle proteine possa essere anche la conseguenza di un rilassamento dei vincoli, la corrispondenza di livelli più alti di sostituzioni di amminoacidi nelle proteine che giocano un ruolo nella riproduzione e nell'immunità sembra essere determinata dalla biologia e dalla selezione.
Le stime statistiche dell'evoluzione adattativa basate sulla genetica delle popolazioni e sulle sequenze proteiche rendono necessari tre avvertimenti a proposito di quanto ci dicono circa l'evoluzione umana. Primo, non esistono di solito dati funzionali diretti che provino o dimostrino se una proteina umana sia davvero differente da un punto di vista funzionale da un ortologo di scimmia antropomorfa. Secondo, le proteine per le quali si sono individuati segni di selezione di solito non incidono sullo sviluppo. Terzo, il proteoma fa solo parte del quadro d'insieme dell'evoluzione genomica. Le sequenze non codificanti, compresi gli elementi cis-regolatori della trascrizione, le regioni non tradotte degli RNA messaggeri, e i segnali di splicing dell'RNA, contribuiscono considerevolmente all'evoluzione, incidendo sul tempo, il luogo e il livello dell'espressione genica. Già a partire dalla pionieristica analisi comparativa della divergenza nella sequenza proteica tra scimmie antropomorfe ed esseri umani, quasi tre decenni fa, ci si è convinti in generale che nell'evoluzione morfologica e comportamentale degli Ominini i cambiamenti nella regolazione genica siano un forza propulsiva più importante rispetto all'evoluzione della sequenza codificante.
L'evoluzione dell'espressione genica umana
Quanto è grande il compartimento funzionale delle sequenze non codificanti l'altro 98% del genoma? Una stima recente indica che forse una quantità due volte maggiore di DNA non codificante è sottoposta a selezione rispetto a quanto avviene per il DNA codificante. Perciò, ci aspetteremmo anche un grande numero di sostituzioni nella discendenza umana, dell'ordine di parecchie centinaia di migliaia, con potenziali conseguenze funzionali nel DNA non codificante. Anche se si fa una stima molto più piccola e più cauta del numero di sostituzioni adattive nel DNA non codificante, per esempio il 2%, si arriva ancora a una cifra di più di 10.000 sostituzioni adattive nei geni umani e nelle loro regioni regolatorie. Il problema è che le sequenze regolatorie sono più difficili da analizzare: non esistono algoritmi che possano dedurre la funzione biologica dai tratti della sequenza intergenica o intronica, per non parlare della possibilità di decifrare come i cambiamenti delle coppie di basi incidano sulla funzione. È pertanto comprensibile il motivo per cui le regioni non codificanti abbiano ricevuto poca attenzione a livello della genetica delle popolazioni. Tuttavia, un numero crescente di studi in genetica quantitativa e sull'evoluzione dello sviluppo ha mostrato che le sequenze regolatorie sono centrali per i cambiamenti nell'espressione e nella morfologia geniche. Si sono rese necessarie nuove metodologie per scoprire l'evoluzione dell'espressione genica e delle sequenze regolatorie.
Un primo passo in avanti verso l'identificazione degli schemi dell'espressione genica specifica dell'uomo è stato fatto di recente utilizzando microarray genomici per analizzare le differenze intraspecie e interspecie nell'espressione genica dei Primati. L'analisi dei profili di espressione dell'RNA provenienti dal lobo prefrontale sinistro (area 9 di Brodmann, che si pensa sia coinvolta nelle funzioni cognitive) di maschi adulti di uomo, scimpanzé e di un orangutan, e provenienti dalla neocorteccia di esseri umani, scimpanzé e macachi, hanno indicato un'apparente accelerazione delle differenze di espressione genica nel cervello umano rispetto ad altri Primati e ad altri tessuti. Le analisi di espressione proteica concordavano inoltre con l'idea che i cambiamenti relativi nei livelli di espressione proteica risultassero accelerati nell'evoluzione del cervello umano e potessero essere rilevati per il 30% circa delle proteine esaminate. La corrispondenza tra i dati relativi all'RNA e quelli relativi al livello proteico indica che i cambiamenti regolatori hanno avuto luogo in un notevole numero di geni. Anzi, un recente esame di esseri umani, eterozigoti in corrispondenza di 13 loci, ha rivelato una variazione allelica nei livelli di espressione genica in corrispondenza di 6 loci. Sia le variazioni intraspecifiche sia la divergenza interspecifica nell'espressione genica sono probabilmente dovute a sostituzioni nelle regioni non codificanti che influenzano la trascrizione o l'abbondanza proteica attraverso meccanismi trascrizionali o post-trascrizionali. Questi dati suggeriscono inoltre che i cambiamenti quantitativi nell'espressione genica non sono altro che una caratteristica generale che accompagna la divergenza tra specie, e che la materia prima per i cambiamenti evolutivi nell'espressione genica sembra ampiamente disponibile nel DNA non codificante.
Gli esperimenti con i microarray propongono molte sfide per gli sviluppi futuri. In particolare, come si possono selezionare i cambiamenti che contribuiscono all'anatomia, alla fisiologia o al comportamento umani rispetto a quelli che non contribuiscono? I dati sull'espressione genica sono correlativi, non definitivi per ciò che riguarda l'identificazione di causa ed effetto. Sono molti i meccanismi evolutivi e genetici che potrebbero contribuire allo schema complessivo che si è osservato. Nel caso di un cambiamento nella composizione di un tessuto (per es., le proporzioni relative dei tipi cellulari), questo sarà accompagnato da profili di espressione alterati, ma molte di queste variazioni saranno una conseguenza indiretta di un mutamento evolutivo, non la causa. Analogamente, i cambiamenti nei livelli o nelle attività delle proteine regolatorie potrebbero influenzare la serie di geni a valle, ma di nuovo sono indiretti e non comportano necessariamente delle sostituzioni in corrispondenza dei loci la cui espressione cambia. Perciò, si debbono intraprendere approcci differenti per identificare i cambiamenti primari nei percorsi di regolazione.
Geni candidati nell'evoluzione umana
Il traguardo finale delle analisi di microarray, della genetica dei tratti quantitativi, della genetica delle popolazioni o di altri metodi di genetica comparativa è l'identificazione di quei geni che sono candidati a essere associati in modo causale alla divergenza fenotipica. Sebbene gli esami su larga scala nell'intero genoma forniscano una panoramica generale, gli esperimenti rigorosi della causalità richiedono un approccio gene per gene. Nella scelta dei geni da studiare più in dettaglio, i genetisti molecolari tenderanno a essere opportunistici, concentrandosi su quei loci per i quali informazioni aggiuntive provenienti dalla biologia umana o da quella degli animali modello suggeriscono un'associazione con un tratto distintivo di maggiore interesse evolutivo, come lo sviluppo craniodentale. Perciò, è improbabile che si ricerchino tutti i tratti distintivi con eguale vigore o successo.
Per dimostrare che un gene è coinvolto nell'evoluzione umana, è necessario valutare due tipi di dati. Primo, sono necessarie le prove funzionali che un gene sia coinvolto in un tratto evolutivo, comportamentale o fisiologico, per formulare delle ipotesi circa il ruolo di un singolo gene. Ciò può derivare dall'analisi delle mutazioni umane in corrispondenza di un locus. Secondo, è necessario analizzare l'evoluzione molecolare e la genetica delle popolazioni del locus per cercare la prova della selezione naturale. Il confronto tra ortologhi provenienti da scimpanzé e altri Primati e Mammiferi, e le analisi della variazione intraspecifica negli esseri umani, possono rivelare segni di selezione positiva al livello della sequenza o di una recente 'spazzata selettiva' attraverso un locus (Tav. I). Prove della selezione positiva sono state rintracciate in corrispondenza di parecchi loci umani. Sebbene questi potrebbero essere fisiologicamente importanti (per es., nell'immunità o nella riproduzione), si ritiene che la maggior parte dei geni studiati sinora non contribuiscano alla divergenza dei tratti morfologici o comportamentali. Più recentemente, si è rivolta l'attenzione a geni candidati identificati a partire da mutazioni umane che influenzano tali tratti caratteristici.
L'evoluzione di un gene che influisce sul linguaggio
La genetica medica umana ha fatto progressi notevoli, e le sofisticate tecniche di mappatura per i polimorfismi stanno accelerando la caratterizzazione dei geni coinvolti nei tratti complessi, in particolare quelli di interesse medico. Uno degli studi più interessanti degli ultimi tempi ha portato all'identificazione del gene FOXP2, le mutazioni del quale sono associate a un disturbo della parola e del linguaggio. Il gene codifica un fattore di trascrizione e pertanto si ritiene che controlli l'espressione di altri geni. L'interesse che circonda FOXP2 deriva dall'osservazione che individui che ne sono affetti sembrano presentare non un danno evidente ma una lesione ai circuiti neurali che interessano i processi del linguaggio.
Si tratta di un nuovo gene umano 'del linguaggio'? No, il gene si trova in altre specie. Infatti, la proteina umana FOXP2 differisce dalla sequenza del gorilla e dello scimpanzé in appena due residui, e dalle sequenze dell'orangutan e del topo in tre e quattro residui rispettivamente. Questa storia è tipica dei geni umani e di altre specie, per il fatto che la maggior parte dei geni hanno ortologhi in altri Mammiferi e animali. Tuttavia, c'è la possibilità che le due sostituzioni nella proteina FOXP2 che evolsero nella discendenza umana siano di importanza funzionale per l'origine del linguaggio.
Per verificare se il gene FOXP2 sia stato il bersaglio della selezione nel corso dell'evoluzione umana, è stato compiuto un esame dettagliato della variazione del nucleotide per una sottoregione di 14 chilobasi (kb) del grande locus FOXP2, del polimorfismo degli amminoacidi in un segmento della proteina e delle sequenze dello scimpanzé e dell'orangutan. Si trovò un insolito eccesso di rari alleli in corrispondenza del locus di FOXP2 umano, e di alleli ad alta frequenza. La ridotta variazione genetica nelle regioni neutre collegate è la conseguenza prevista di una spazzata selettiva, perciò queste osservazioni concordano con l'azione della selezione naturale sul locus di FOXP2. Stime del tempo di fissazione delle due sostituzioni di amminoacidi le collocano nell'arco degli ultimi 200.000 anni dell'evoluzione umana, una correlazione intrigante con l'età stimata di H. sapiens.
Tuttavia, si dovrebbe notare che non ci sono dati biologici a sostegno dell'ipotesi che queste sostituzioni di amminoacidi sono funzionalmente importanti. Nella regione di 14 kb esaminata, esistono più di 100 differenze fisse; l'intero locus di FOXP2 è esteso (267 kb), e si ritiene esistano più di 2000 differenze tra i geni di FOXP2 degli esseri umani e degli scimpanzé. Non è stata fatta alcuna valutazione delle potenziali sequenze regolatorie non codificanti che potrebbero aver contribuito alla divergenza nel ruolo di FOXP2 negli Ominidi. Le differenze dei tratti distintivi sono spesso dovute a cambiamenti nelle reti regolatorie che governano lo sviluppo, e non è necessario siano in regioni codificanti (anche se ciò sarebbe molto più conveniente, dati gli appena due cambiamenti nella proteina FOXP2 dell'uomo). Poiché FOXP2 è un fattore di trascrizione, i cambiamenti nell'espressione di FOXP2 potrebbero essere di importanza funzionale ed evolutiva.
La tipica architettura genetica che sta alla base dei tratti complessi rende estremamente improbabile che FOXP2 fosse l'unico gene sottoposto a selezione nell'evoluzione delle nostre capacità linguistiche. Tuttavia, non abbiamo modo di valutare il contributo relativo di FOXP2 e di altri geni candidati. La lezione incoraggiante di FOXP2 è che la genetica medica ha fornito una interessante chiave di lettura per una rete regolatoria che influenza lo sviluppo della capacità linguistica. Studi ulteriori di FOXP2 dovrebbero portare, come minimo, a una migliore comprensione della biologia neuroevolutiva della parola e del linguaggio, e forse alla scoperta di più geni con interessanti storie evolutive.
Le funzioni dei geni selezionati
I tre approcci su scala genomica qui evidenziati ‒ genetica delle popolazioni, genomica comparativa e studio dell'espressione genica ‒ sono tutti riusciti a trovare ciò che stavano cercando: migliaia di potenziali sostituzioni adattative codificanti, differenze regolatorie nell'espressione genica e duplicazioni e riarrangiamenti genici. Ciascuno ha prodotto molti candidati da vagliare scrupolosamente e, curiosamente, a causa dei diversi sistemi di ricerca utilizzati, non c'è praticamente nessuna sovrapposizione nelle serie di cambiamenti genetici che sono stati esaminati. È quasi certo che, come in altre discendenze, tutti questi tipi di meccanismi genetici hanno contribuito all'evoluzione degli Ominidi. La sfida cruciale adesso è quella di ottenere dei dati funzionali per i singoli geni e di esaminare minuziosamente l'evoluzione molecolare dei candidati in cerca di segni di selezione.
Per collocare un qualsiasi gene candidato in un contesto funzionale dell'evoluzione dei tratti umani, saranno essenziali progressi nella neurobiologia evolutiva dei Primati e dell'uomo. I non Primati hanno un valore limitato come modelli dello sviluppo e della funzione della neocorteccia nei Primati e negli Ominidi, e pure come modelli della funzione di proteine come FOXP2 nello sviluppo e nell'elaborazione delle reti neurali.
Il lavoro empirico diretto sui Primati in fase di sviluppo, che si trova ad affrontare seri limiti metodologici così come autentiche questioni etiche, sarà sempre più necessario se si vuole andare oltre le associazioni e le correlazioni. Esaminare il ruolo funzionale di quelli che potrebbero essere cambiamenti impercettibili negli ortologhi umani dei geni dei primati, un compito sconfortante nella maggior parte delle specie modello tecnicamente sviluppate, sarà ancora più difficile.
Esistono due modi diretti per accrescere il potere della genetica dell'evoluzione umana. In primo luogo, si potrebbe incrementare il valore della genomica comparativa scimpanzé-uomo stabilendo la sequenza del genoma del gorilla, che rappresenta la scimmia antropomorfa più prossima che si è separata dagli esseri umani e dagli scimpanzé. Ciò ci aiuterebbe a determinare la polarità dei cambiamenti genetici distinguendo i cambiamenti presenti nella discendenza umana da quelli presenti nella discendenza degli scimpanzé. In secondo luogo, 6×109 esseri umani incrociantisi tra loro rappresentano una risorsa molto ampia per l'identificazione di mutazioni rare (per es., nel FOXP2) con impercettibili effetti comportamentali o evolutivi, e per la mappatura della variazione genetica che sta alla base della variazione morfologica; entrambe le cose potrebbero portare all'individuazione dei geni che governano la formazione dei tratti distintivi dell'uomo e che potrebbero aver avuto un ruolo nell'evoluzione degli Ominidi.
Nonostante la nostra maggiore comprensione dell'architettura genetica funzionale, resta la tendenza ad associare lo sviluppo, la funzione o l'evoluzione di un tratto con i singoli geni (geni 'per' la parola, il cancro, ecc.). Il fantasma del 'mostro di belle speranze' ancora infesta la biologia ed è, sfortunatamente, un errore molto frequente nelle pubblicazioni scientifiche e non specialistiche. Forse anche il velleitarismo è una parte sostanziale della natura umana, ma sembra improbabile che i tratti che ci interessano di più ‒ il bipedismo, la morfologia scheletrica, la morfologia craniofacciale, le dimensioni cerebrali e la parola ‒ siano il prodotto della selezione di pochi geni di maggior rilievo. Proprio come la paleoantropologia adesso riconosce l'esistenza di un complesso schema di filogenesi degli Ominini e ammette alcune incertezze nell'identificazione degli antenati comuni a lungo cercati, e la neurobiologia comparativa va alla ricerca di spiegazioni più accurate per giustificare le capacità umane, le lezioni della genetica dei sistemi modello e della genomica comparativa dovrebbero farci giungere alla conclusione che la genetica dell'evoluzione dei tratti distintivi degli Ominidi è, in realtà, impercettibile e complicata.
Non sottolineare questo, e abbandonarsi a facili sensazionalismi infondati, offre inutilmente il fianco a chi nega la realtà dell'evoluzione.
Bibliografia
Carroll 2001: Carroll, Sean B. - Grenier, Jennifer K. - Weatherbee, Scott D., From DNA to diversity: molecular genetics and the evolution of animal design, Oxford, Blackwell Scientific, 2001.
Davidson 2001: Davidson, Eric H., Genomic regulatory systems: development and evolution, San Diego-London, Academic Press, 2001.
Enard 2002: Enard, Wolfgang e altri, Intra- and interspecific variation in primate gene expression patterns, "Science", 296, 2002, pp. 340-343.
Enard 2002: Enard, Wolfgang e altri, Molecular evolution of FOXP2, a gene involved in speech and language, "Nature", 418, 2002, pp. 869-872.
Gould 1977: Gould, Stephen J., Ontogeny and phylogeny, Cambridge (Mass.)-London, Belknap Press, 1977.
Klein, Takahata 2002: Klein, Jan - Takahata, Naoyuki, Where do we come from? The molecular evidence for human descent, New York-Berlin, Springer, 2002.
Moggi-Cecchi 2001: Moggi-Cecchi, Jacopo, Human evolution: questions of growth, "Nature", 414, 2001, pp. 595-597.
Nachman 2001: Nachman, Michael W., Single nucleotide polymorphisms and recombination rate in humans, "Trends in genetics", 17, 2001, pp. 481-485.
Ohta 2002: Ohta, Tomoko, Near-neutrality in evolution of genes and gene regulation, "Proceedings of the National Academy of Sciences USA", 99, 2002, pp. 16134-16137.
Relethford 2001: Relethford, John H., Genetics and the search for modern human origins, New York-Chichester, Wiley-Liss, 2001.
Rice 2002: Rice, Sean H., The role of heterochrony in primate brain evolution, in: Human evolution through developmental change, edited by Nancy Minugh-Purvis, Kenneth J. McNamara, Baltimore-London, Johns Hopkins University Press, 2002, pp. 154-170.
Ruff 1997: Ruff, Christopher B. - Trinkhaus, Erik - Holliday, Trenton W., Body mass and encephalization in Pleistocene Homo, "Nature", 387, 1997, pp. 173-176.
Sabeti 2002: Sabeti, Pardis C. e altri, Detecting recent positive selection in the human genome from haplotype structure, "Nature", 419, 2002, pp. 832-837.
Wilkins 2002: Wilkins, Adam S., The evolution of developmental pathways, Sunderland (Mass.), Sinauer Associates, 2002.
Wood, Collard 1999: Wood, Bernard - Collard, Mark, The human genus, "Science", 284, 1999, pp. 65-71.
Yan 2002: Yan, Hai e altri, Allelic variation in human gene expression, "Science", 297, 2002, pp. 1143.
Tavola I
Selective sweeps
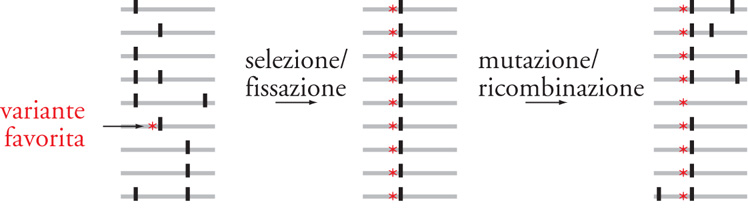
Se un cambiamento in un gene è adattativamente positivo, allora la selezione potrebbe condurre alla fissazione dell’allele che porta quel cambiamento (a sinistra e al centro della figura). Nel processo, la variazione neutra in corrispondenza di siti collegati viaggia insieme ai siti selezionati (il cosiddetto ‘autostop genetico’); ciò è noto come selective sweep, ossia una sorta di ‘spazzata selettiva’. I limiti fisici dello sweep dipendono dalla forza del linkage statistico tra siti selezionati e siti adiacenti. Dopo uno sweep, la variazione potrebbe iniziare nuovamente ad accumularsi, e inizialmente vi sarà una frequenza relativamente alta di polimorfismi rari (a destra della figura). Viene utilizzato un dato statistico (D) che misura la neutralità selettiva. Se le frequenze dei polimorfismi sono distorte, con un eccesso di alleli rari, ciò dà un valore negativo (valore neutro uguale a 0) e può essere indicativo di una recente selective sweep. D è sensibile ad altri fattori a parte la selezione che possono anche produrre un valore negativo. Una espansione recente nella dimensione di una popolazione a partire da una popolazione relativamente piccola produrrà schemi simili di variazione genetica e di valori di D. Nelle popolazioni umane, la storia della popolazione (per es., impulso ed espansione) e la struttura della popolazione (etnicità, migrazione e immigrazione) influenzeranno i valori di D in corrispondenza di tutti i loci, mentre la selezione influenzerà i valori di D in corrispondenza dei loci selezionati e collegati. I valori medi di D per 437 loci vanno da -0,69 a -1,25, a seconda dei metodi di campionatura, a indicare che la struttura e la storia della popolazione hanno lasciato un segno. Questi valori negativi sottolineano una sfida nella genetica dell’evoluzione umana per distinguere le selective sweeps in corrispondenza dei loci dagli effetti che hanno sede nella popolazione. Il valore di D ottenuto per il locus di FOXP2 umano era -2,20, il secondo più grande valore negativo tra tutti i geni umani sinora sopravvissuti. Altri metodi sono stati sviluppati perscoprire la selezione positiva: per esempio, identificando le aree di estesa omozigosità degli aplotipi. È importante sottolineare che tutte queste metodologie rilevano segni di recente selezione nella discendenza di Homo sapiens. I precedenti 5-7 milioni di anni della storia genetica degli Ominini, un periodo in cui sappiamo dalle testimonianze fossili che sono sorte molte caratteristiche umane, non sono presi in considerazione da questi metodi.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


