Entomologia
Enciclopedia del Novecento (1977)
Entomologia
di Baccio Baccetti
Entomologia
sommario: 1. Introduzione. 2. I pionieri. 3. Il sistema degli Insetti, la faunistica e la morfologia del dermascheletro. 4. La micromorfologia. 5. La composizione chimica degli Insetti. 6. La fisiologia degli Insetti. 7. L'etologia entomologica. 8. Genetica e dinamica di popolazione. 9. L'entomologia economica. a) Gli Insetti merceologicamente utili. b) Insetti e agricoltura. c) Insetti e foreste. d) Insetti e suolo. e) Insetti e virus. f) Insetti e Vertebrati. 10. Il problema della lotta contro gli Insetti. 11. Il rilancio dell'entomologia. 12. Le nuove aperture della sistematica. □ Bibliografia.
1. Introduzione
L'entomologia è la parte della zoologia che si occupa dello studio degli Insetti o Esapodi. Il progredire delle cognizioni che la compongono, per certi aspetti parallelo a quello della zoologia, è stato anche regolato e stimolato da esigenze particolari che furono prima di ordine economico e medico, poi a carattere di hobby, infine di studio biologico puro. La classe degli Insetti comprende quasi un milione di specie descritte, e cioè circa i tre quarti dell'intero regno animale, ma si può prevedere che il numero delle specie realmente esistenti sia almeno tre o quattro volte superiore e che il numero d'individui che pullulano sul globo sia, praticamente, incalcolabile: tutto ciò significa che questa classe è caratterizzata da un grado estremo di plasticità e adattabilità a tutti gli ambienti, tale da condurla a frazionarsi in un numero sterminato di forme. L'uomo, nella sua progressiva conquista di tutti gli spazi del pianeta, si è trovato inevitabilmente a cozzare contro questa popolazione che abitava il mondo già da lungo tempo prima di lui (i primi Insetti sono comparsi 350-400 milioni di anni fa) ed ha dovuto studiare a fondo le tecniche per difendersi da essa. Tanto per fare un esempio, circa il 12-15% della produzione agricola mondiale viene ogni anno distrutto dagli Insetti, e i soli prodotti immagazzinati che questi distruggono potrebbero sfamare 130 milioni di persone. È superfluo poi ricordare l'enorme numero di malattie che gli Insetti trasmettono all'uomo e agli animali domestici. Il primo impulso che spinse l'uomo allo studio dell'entomologia fu perciò essenzialmente di ordine economico; poi, la necessità di riconoscere, denominare e classificare lo sterminato numero di specie determinò l'avvento della sistematica, e questa a sua volta fu seguita dalla formazione di un esercito di collezionisti amatori; infine divenne ben chiaro che l'organismo-tipo dell'Insetto costituisce un eccellente modello biologico e su di esso s'impostarono numerose discipline. La genetica, l'etologia, l'endocrinologia, l'ecologia affondarono le loro radici proprio nello studio degli Insetti, e al tempo stesso lo vivificarono rapidamente, suggerendo sempre nuove soluzioni per risolvere i problemi medico-economici. Lo studio degli Insetti, poiché questi rappresentano una tra le più specializzate classi del regno animale, ha sempre richiesto attitudini e forma mentis particolari, e per alcuni aspetti il suo progredire è stato quindi indipendente da quello del complesso delle altre discipline biologiche: talora infatti ha registrato un anticipo, tal altra un ritardo, e l'entomologo è stato considerato di volta in volta un pioniere o un sorpassato, ma in ogni caso un individuo fuori moda, pieno di fantasia e d'illusioni, solitario, bastante a se stesso, facile alla presunzione e perciò al litigio.
2. I pionieri
Non è facile individuare il momento in cui l'entomologia è nata, nè poter dire che cosa abbia realmente dato all'umanità, che cosa rappresenti ora, se avrà una ragion d'essere in futuro. Si suole assumere il 1602 come anno di nascita di questa disciplina (v. Goidanich, 1957; v. Montalenti, 1960; v. Baccetti, 1965), data di pubblicazione della famosa opera di U. Aldrovandi, De animalibus insectis libri septem: prima di allora si era fatto assai poco. Aristotele per primo aveva individuato negli Insetti un particolare gruppo di animali senza sangue, distinti da Molluschi e Crostacei, ne aveva elencato alcune specie e aveva indicato nell'apicoltura un primo argomento d'interesse pratico. Bachi da seta, introdotti in Europa nel Medioevo, e api, molto empiricamente studiati, continuarono poi a essere l'argomento entomologico principe per molti secoli, con ben pochi progressi scientifici. U. Aldrovandi (1522-1605), naturalista bolognese, pubblicò ottantenne un grosso trattato sugli Insetti, compendiandovi una quantità di osservazioni, accumulate in molti decenni, molte delle quali di prima mano. L'entomologia era allora tutta da scoprire e molte idee originate da premesse teoriche e da concezioni completamente errate, come quella della generazione spontanea, dovevano essere radicalmente mutate. Sarà questa la via che percorreranno i grandi entomologi del Seicento: il biologo F. Redi (1626-1698), che chiarirà una volta per tutte il mistero della metamorfosi degli Insetti; il microscopista M. Malpighi (1627-1694), fondatore dell'anatomia degli Insetti e primo dissettore e illustratore dei loro organi interni ed esterni; e 3. Swammerdam (1637-1680), che metterà a confronto dati biologici, morfologici e fisiologici, e comincerà a delineare le soluzioni dei massimi problemi entomologici, quali la sessualità delle tre caste delle api, l'emimetabolia consistente in un ciclo vitale dapprima in acqua e poi in aria con le conseguenti differenti morfologie specializzate in larve e adulti delle Efemere, le peculiarità dell'anatomia del pidocchio. Ancora, fra molti errori, l'aristotelico padre F. Buonanni descrive nel 1691 le morfologie di molti apparati boccali pungenti e succhianti (Afanitteri, Mallofagi, Ditteri, Lepidotteri, Tisanotteri, Rincoti); e, in un susseguirsi di scoperte, A. Vallisnieri (1661-1730) mette in luce numerosissime etologie, quali quelle dell'Empusa (1715), dei vari Ditteri parassiti degli animali domestici (1713), dei Tentredinidi delle rose (1713), del Crisomelide del giglio (1713).
A. Leeuwenhoek (1632-1723) scopre nel 1695 la partenogenesi e la ovoviviparità degli Afidi e descrive in molti insetti (1695-1722) accoppiamento e spermatozoi, attribuendo a questi ultimi la capacità fecondatrice. I vari tipi di metamorfosi sono indagati da I. Goedart (1662), che vi dedica un libro con stravaganti illustrazioni, e da J. Ray (1710), mentre R. A. Réaumur (1683-1757), in un'opera in sei volumi stampata per la prima volta a Parigi fra il 1734 e il 1742, si dedica a fondo allo studio comparato dei costumi degli Insetti, elaborando il primo trattato di etologia comparata. Intanto, i grandi viaggiatori incominciano a portare a conoscenza degli scienziati europei strane faune esotiche ed a suscitare in loro il desiderio e il gusto di illustrarle: fra i primi a sentirne il fascino furono G. MacGraw de Liebstad (1648), che descrisse e raffigurò fra gli animali brasiliani meravigliosi Fasmoidei, Scarabeidi, Rincoti, e M.S. Merian che nel 1705 illustrò gli Insetti del Suriname. Successivamente, E. Albin (1720) si dedicò all'entomofauna inglese, illustrandone i Lepidotteri e pochi Tentredinidi, Coleotteri, Ditteri e Neurotteri. Tutta questa messe di conoscenze rese necessaria l'istituzione di un sistema classificatorio: chiariti nelle linee essenziali i vari tipi di metamorfosi, la particolare anatomia e morfologia dei differenziatissimi adulti, la loro etologia, gli scienziati si trovarono di fronte alla necessità di dare un nome all'enorme quantità di specie che via via andavano enumerando.
Tale esigenza si rese manifesta in un paio di splendidi trattati usciti alla metà del Settecento: imponente fu l'opera pubblicata fra il 1746 e il 1761, con finissime miniature, da A. J. Roesel, Insekten Belustigung, in cinque tomi ove erano condensate e illustrate tutte le cognizioni entomologiche dell'epoca e un'infinità di notazioni sulle faune esotiche. Altrettanto importante, anche se più sintetico, fu il trattato di C. De Geer (1752), maggiormente incline alla morfologia comparata. Questi autori ebbero sottomano splendide raccolte di Insetti, poiché, proprio in quell'epoca, iniziò a diffondersi il fenomeno del collezionismo, durato poi per tutto il Settecento e anche per tutto il secolo seguente, soprattutto per merito di dilettanti. Occorreva però una chiave per potersi orientare nella classificazione della straordinaria moltitudine di forme, e questa chiave fu offerta da C. Linneo che nel suo trattato Systema naturae (di cui vennero stampate tredici edizioni, dal 1735 al 1788-1793) introdusse la nomenclatura binomia e incasellò tutti gli esseri viventi in specie, generi, ordini, classi. La data fatidica che segna l'inizio dell'entomologia sistematica è il 1758, anno di pubblicazione di quella famosa editio decima linneana che realmente fa testo: da allora i singoli ordini di Insetti vengono arricchiti di specie, e ancora numerosi altri ne vengono istituiti a opera di una moltitudine di entomologi che, armati di retino, bottiglie, spilli e scatole, immortalano il proprio nome legandolo alla descrizione di categorie sistematiche nuove, nell'ambito di quello che è il gruppo di animali più popoloso del globo. L'entomologia, fino allora dominio dei più abili microscopisti e dei più geniali biologi, tutti professionisti delle massime scuole mediche dell'epoca, che avevano saputo compiere mirabili scoperte di valore zoologico e biologico generale, dopo Linneo e per più di un secolo diviene proprietà degli amatori, i quali intraprendono la pubblicazione delle faune regionali.
3. Il sistema degli Insetti, la faunistica e la morfologia del dermascheletro
Dalla metà del Settecento a tutto il secolo scorso si assiste all'edificazione del sistema degli Insetti, che si compone e completa man mano che l'esplorazione di nuove regioni svela l'esistenza di forme nuove. Direttamente dallo studio delle collezioni gli entomologi formulano le diagnosi di nuove entità e compongono un quadro sistematico essenzialmente fondato sulla morfologia del dermascheletro. L'approccio, anche se casuale (il dermascheletro è la parte del corpo degli Insetti più facile da esaminare, più solida, più colorata e la sola che si conservi nelle collezioni di esemplari spillati), non è fallace, in quanto è proprio il dermascheletro l'organo che più strettamente interagisce con l'ambiente, che va incontro a evoluzione, che presenta i più vivaci adattamenti morfologici e i più esemplari mimetismi, che infine nel corso della vita necessita di modificazioni legate ai cambiamenti di abitudini fra giovani (larve) e adulti; esso, ovviamente, non può riassumere l'intera essenza di una specie, ma ne è generalmente specchio fedele. I faunisti, che descrivono la morfologia esterna degli esemplari catturati ed edificano la sistematica con il metodo linneano, rappresentano quindi la massima espressione del lavoro entomologico fino a tutto l'Ottocento.
Allo studio formale dell'aspetto esteriore dell'Insetto è legato il sorgere della scienza iconografica entomologica. Dopo le prime rozze (ma, in quanto caricaturali, efficacissime) incisioni in legno di T. Mouffet, di A. Mattioli, di A. Vallisnieri, nel Seicento il ritratto dell'Insetto andò progressivamente raffinandosi e finalmente nel secolo successivo si affermò a opera di una serie affascinante di impeccabili miniaturisti: P. Cramer, C. Stoll, J. Voets, D. Drury, E. Donovan ecc., degni successori dei vari Sano di Pietro, Liberale da Verona, che nel Medioevo avevano illustrato i graduali con fantasiose farfalle e libellule, segnando una pagina indimenticabile nella storia dell'arte mondiale. Nell'Ottocento si impone il disegno a inchiostro di china, sempre semischematico, il più adatto a esaltare i duri spigoli della cuticola sderificata: i suoi cultori in Italia saranno di primissimo ordine e le maggiori scuole, da quella romana di B. Grassi, a quella napoletana fondata da A. Berlese e proseguita da F. Silvestri, a quella genovese di R. Gestro, produrranno eccellenti specialisti. Il più grande di tutti i tempi sarà A. Berlese che, scienziato e artigiano a un tempo, nelle soffitte dell'antico palazzo ospitante la Facoltà di Agraria di Napoli prima e in quelle del palazzo che ospiterà la Stazione Entomologica di Firenze poi, con attività indefessa tirerà da solo disegni, zinchi, rami di uno sterminato numero di Artropodi, sui quali edificherà il più esaltante trattato di entomologia che sia mai stato scritto.
Il primo sistema degli Insetti modernamente impostato (basato sulla presenza e sulla forma delle ali) è quello del Linneo, che è il seguente:
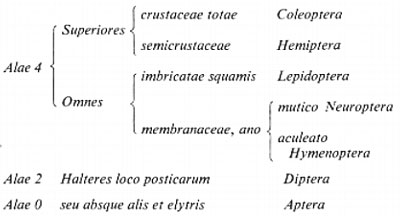
Dopo secoli di ampliamenti, possiamo ritenere soddisfacente lo schema che riportiamo nella pagina seguente.
Questa classificazione non può certo ritenersi definitiva: il progredire delle conoscenze ha consentito di individuare numerosi altri caratteri dell'Insetto, oltre alla morfologia dermascheletrica, così che l'anatomia prima, poi via via le formule cromosomiche, gli spermatozoi e le uova, gli amminoacidi dell'emolinfa, le affinità sierologiche hanno offerto sempre nuovi elementi su cui fondare la filogenesi e con essa una classificazione imperniata sullo studio del cammino percorso dall'evoluzione. Il collezionismo, che non è in decadenza, anzi conosce ora livelli mai raggiunti prima, offre grandi serie di esemplari conservati attualmente indispensabili per lo studio della variabilità, fenomeno che la genetica ha posto in posizione di preminenza agli occhi dei sistematici: la crisi investe soltanto il classico concetto di specie caro ai collezionisti ottocenteschi, per cui non si cercano più diagnosi discriminatorie, ma anzi, valutando una specie in tutto il suo areale, si tende a esaltarne la frammentazione in razze, se ne cercano i punti di contatto con le specie vicine. L'amatore è sempre più in difficoltà, le grandi serie complicano il quadro; il numero delle specie ritenute valide va riducendosi e molte entità vengono ridimensionate e spostate a livello di razza. Dopo la genetica, che ha attratto l'attenzione degli entomologi sulla microevoluzione, un'altra disciplina vivifica le collezioni: compare infatti la biogeografia, che cerca di svelare il perché dei popolamenti e delle differenziazioni faunistiche delle più svariate regioni. L'opera pionieristica di R. Jeannel addita suggestive correnti di popolamento, basate su connessioni territoriali ora perdute, mentre. E. Mayr (v., 1963) individua nei fattori geografici la principale causa dell'affermarsi di specie e razze animali nuove. Su queste premesse fioriscono le interpretazioni sui tre tipi di faune entomologiche più importanti: quelle insulari, quelle di alta quota e quelle di caverna.
La geologia offre gli elementi critici necessari per valutare l'attendibilità di molte delle ipotesi avanzate. Dopo un periodo di crisi, durante il quale la diffusione accidentale (anche meteorica) sembra responsabile degli spostamenti di piccoli campioni di faune (le ‛propagule') da una terra all'altra (v. MacArthur e Wilson, 1963), attualmente il discorso si fa più equilibrato e verte essenzialmente sulla individuazione di quelle faune che più difficilmente subiscono un trasporto passivo. Molti Coleotteri, Ortotteri, Atterigoti, scarsi volatori, con esigenze ecologiche precise, offrono al biogeografo gli elementi chiave (v. MacKerras, 1970) per postulare ponti territoriali.
Ovviamente il biogeografo deve essere pur sempre uno specialista nella sistematica del gruppo che intende utilizzare e del quale può meglio di ogni altro vagliare le capacità di spostamento: difatti, attualmente, i più convinti sostenitori della teoria delle propagule si incontrano fra gli specialisti di forme che fanno parte del ‛plancton aereo', mentre i più grandi fautori dei ponti territoriali si trovano fra gli studiosi dei Coleotteri, degli Ortotteri, ecc.

Nello schema di classificazione riportato nella tab. I, a pag. 636, appaiono evidenti le caratteristiche morfologiche fondamentali di tutta la classe degli Esapodi, come pure quelle che differenziano i vari gruppi che la costituiscono, caratteristiche che possono essere comuni a tutti gli stadi che l'Insetto attraversa prima di diventare adulto, o che invece possono riguardare solo alcuni degli stadi e non altri. Queste caratteristiche riguardano la forma presentata da una parete compatta e dura disposta sopra lo strato periferico di cellule (epidermide) dell'organismo degli Insetti, e precisamente la porzione rivolta verso l'esterno del suo glycocalyx, enormemente arricchita e complicata, denominata cuticola. Tutte le cellule sono normalmente rivestite da una guaina glicoproteica (appunto il glycocalyx), che usualmente comprende acido sialico e mucine; quelle dell'epidermide degli Insetti presentano esternamente un rivestimento costituito da una mescolanza di proteine strutturali (artropodina, resilina, sderotina) e di un glucide particolare, la chitina, in unione a polifenoli e cere che inglobano i lunghi microvilli proiettati in fuori dalle cellule stesse. Questa complicata corazza, spesso dura e rigida, ha aree più mobili che fungono da cerniere, e aree compatte (sderiti); essa viene periodicamente rinnovata con l'accrescimento dell'Insetto ed è la principale responsabile della sua forma. Le tecniche di studio più recenti, quali la microscopia elettronica a scansione, ne confermano la grande importanza evolutiva e l'utilità a scopo classificatorio. Il suo studio, anche con i più moderni sistemi, procede in modo autonomo rispetto a quello delle strutture cellulari che l'elaborano, ed è e rimane dominio dei sistematici, i quali in esso esauriscono di solito le loro diagnosi. Questa situazione costituisce indubbiamente un grosso limite per l'entomologia attuale. Il livello submicroscopico che lo studio della cuticola tende ora a raggiungere va però colmando la grossa distanza che separava i morfologi dagli anatomisti e rende sempre più probabile un confluire di interessi e perciò la comparsa di una sistematica a più ampio respiro. Lo studio minuto della cuticola allaccia dunque la morfologia del dermascheletro con la scienza delle strutture interne dell'Insetto; cioè con la microanatomia. Il loro insieme si può definire micromorfologia.
4. La micromorfologia
Lo studio minuto della forma degli Insetti ha avuto quattro periodi, corrispondenti a quattro momenti tecnici ben distinti, che potremmo definire il periodo delle dissezioni al microscopio ottico, il periodo dello studio delle sezioni istologiche al microscopio ottico, il periodo della microscopia elettronica a trasmissione, il periodo della microscopia elettronica a scansione.
Il primo periodo, che copre praticamente tutto il Settecento, è legato ai grandi dissettori: M. Malpighi, J. Swammerdam, P. Lyonet, e precisa l'anatomia topografica dell'Insetto. Si scopre che il canale alimentare può dividersi in tre grandi segmenti: intestino anteriore, medio e posteriore, dei quali il primo può ancora ripartirsi in esofago, ingiuvie e ventriglio, mentre il secondo è più uniforme, provvisto di ciechi e separato per mezzo di valvole (cardias e piloro) dall'ultimo tratto, pure distinto in tre regioni (ileo, colon e retto). Malpighi scopre lunghi e convoluti tubi ciechi che sboccano nella regione pilorica: sono i tubi malpighiani, che verranno poi riconosciuti come i principali organi escretori degli Insetti. Swammerdam scopre nel retto delle introflessioni ispessite, che chiama papille rettali. Si descrive il tessuto adiposo interstiziale, l'enorme dimensione delle ghiandole salivari nelle larve tessitrici di bozzoli, la complicatissima muscolatura, la metameria del sistema nervoso, l'intricato sistema tracheale che conduce l'aria ai tessuti.
A questo punto incomincia il secondo periodo: gli entomologi imparano a sezionare microtomicamente e a colorare in modo opportuno il loro materiale, usufruendo delle tecniche che l'istologia dei Vertebrati andava mettendo a punto; aprono così la via all'istologia e citologia entomologica, che praticamente occuperà l'Ottocento e i primi decenni del secolo successivo. Si incomincia a discutere di tessuti connettivi, di un particolare tipo di neurone negli Invertebrati, dell'esistenza di glia e di mielina negli Insetti; si osservano diversi tipi di muscoli (fibrillari e afibrillari, tutti però striati) in tutti gli Insetti volatori; si dimostra che una cuticola simile a quella del dermascheletro è presente entro gli organi con esso collegati (intestino anteriore e posteriore, trachee, ghiandole esocrine), mentre mesenteron e tubi malpighiani hanno all'interno un orletto a spazzola; si riconoscono diversi strati nella cuticola dermascheletrica. La letteratura si fa sempre più ricca e complessa. Chi per primo collega e inquadra l'enorme corpo di nozioni è A. Berlese nel suo già ricordato trattato (1909-1925) rimasto fondamentale nella scienza entomologica.
Con l'affinarsi di questo tipo di studio lo iato fra morfologi esterni e istologi va sempre più approfondendosi e sembra divenire incolmabile quando, verso gli anni quaranta del secolo in corso, si apre il periodo della microscopia elettronica a trasmissione, accolta con estrema circospezione dagli entomologi tradizionali, che temono di vedere in un sol colpo superate tutte le loro tecniche e le loro nozioni. Alcuni istologi però si lanciano sulla nuova via e vi trovano la soluzione a grandi problemi aperti da troppo tempo e ritenuti insolubili. Una valutazione globale del ruolo svolto dalla nuova tecnica sulle conoscenze entomologiche tradizionali è data da G. A. Edwards (v., 1960) e B. Baccetti (v., 1966). La microscopia a luce polarizzata aveva già fornito preliminari ragguagli sulla stratificazione cuticolare per merito di A. Richards (v., 1951), sulla striatura delle fibre muscolari (v. Schmidt, 1937) e sulla presenza di collagene nei tessuti connettivi (v. Baccetti, 1956). Il microscopio elettronico consente di svelare numerosi tipi di muscoli striati, di definire i vari tipi di neurone, di confermare l'esistenza negli Insetti di un collagene simile a quello dei Vertebrati, di delineare la struttura e il funzionamento di tutti gli epiteli assorbenti (vari tratti intestinali), secretori (ghiandole esocrine, endocrine, mesenteron, epidermide), escretori (tubi malpighiani, cellule pericardiali, reni cefalici degli Atterigoti); ancora, fonda le basi della spermatologia, dimostrando negli Insetti un modello di spermatozoo peculiare, evolutosi sul modello primitivo a coda ‛9+2', ma complicatosi, in seguito all'acquisizione della fecondazione interna, con l'aggiunta di 9 microtubuli accessori; inoltre permette un'importante chiarificazione della fine struttura delle uova, i cui gusci elaboratissimi sono costituiti da strati che ricordano quelli della cuticola dermascheletrica, e muniti di particolari aree pervie per la respirazione e la fecondazione. Occhi, sensilli meccanocettori, ‛orecchi' vengono finalmente compresi a livello dello loro morfologia raffinatissima; la cuticola, soprattutto, viene finalmente analizzata a fondo. Tutte le sculture peculiarissime degli Insetti, consistenti in squame, striature, fossette, peli in posizione fissa, capaci di conferire al tegumento caratteri stabili e specifici di sagoma, consistenza e perfino colore, appaiono ora come il risultato dell'azione secretrice di distretti ben precisi di ciascuna cellula epidermica, sollecitata da una particolare spinta ormonale. Sono soprattutto gli studi di M. Locke (v., 1967) che consentono la soluzione di questo problema fondamentale dell'Insetto, animale identificabile dall'esterno per l'aspetto stabile del proprio tegumento.
Sorge così netto il contrasto tra microanatomisti e sistematici: questi ultimi, infatti, aderendo ai concetti della morfologia esterna tradizionale di centinaia di migliaia di entità, subiscono il continuo contrasto tra il desiderio di trovare caratteri nuovi per definire nuove specie e il timore per la potenziale capacità di sconvolgere così il vacillante inquadramento da loro accettato, venendo definiti da alcuni ‛fisiologi' e in un certo senso avvicinati agli etologi, da altri, più seccamente, ‛biofisici'. L'entomologia latina, tradizionalista per eccellenza, è quella che più deve temere da questa situazione. L'avvento della microscopia elettronica a scansione fa però cadere la divergenza, consentendo la visione delle tre dimensioni dermascheletriche a un livello di risoluzione prima impensabile, ma non in un modo diverso da quello dei vecchi sistematici della cuticola: però quest'ultima è sempre vista in collegamento con le cellule epidermiche: i sensilli collegati ai nervi, e le strutture sempre riferite ai caratteri chimico-fisici del dermascheletro. Così, senza traumi, i sistematici possono accedere al mondo delle ultrastrutture.
Il periodo è appena iniziato, ma durerà certo a lungo e appare foriero di grandi acquisizioni, soprattutto per la sistematica dei piccoli organismi (v. Salked e Wilkes, 1968; v. Hinton, 1970; v. Baccetti e Capra, 1970).
5. La composizione chimica degli Insetti
Un secondo approccio alla conoscenza della complessa macchina dell'organismo è quello di indagare come essa funzioni, cioè la ricerca fisiologica. La fisiologia è però scienza biologica di sintesi ‟verso la quale tutte le altre convergono" (v. Bernard, 1865) ed esige quindi la precisa conoscenza morfologica e chimica delle strutture in esame. Si comprende così come le difficoltà delle risoluzioni tecniche di alcuni problemi di istomorfologia, di citochimica e di biochimica abbiano potuto limitare l'espansione della fisiologia entomologica: questa è praticamente nata negli anni trenta a seguito di precise esigenze pratiche (l'agricoltura e la medicina dovevano controllare in modo razionale gli Insetti dannosi) e grazie all'opera geniale e precorritrice di un biologo di Cambridge, V. B. Wigglesworth. Punto di incontro fra strutturistica e biochimica dinamica, in meno di cinquant'anni la fisiologia degli Insetti ha compiuto un ragguardevole cammino ed è oggi una delle branche più valide della biologia generale, tanto che appare difficile finanche la semplice elencazione delle principali acquisizioni da essa realizzate. Le esigenze iniziali furono quelle di capire come funzionasse la cuticola, come venisse rinnovata e modellata e, conseguentemente, quali fossero i meccanismi di base dell'imponente fenomeno della metamorfosi. L'indagine fu successivamente estesa ai fluidi circolanti nell'emocele (altra caratteristica peculiare degli Insetti, che, privi di un vero e proprio sistema circolatorio, hanno tutti gli organi immersi in un'enorme lacuna), quindi al sistema nervoso, alla fibra muscolare, alla secrezione e all'escrezione: le acquisizioni che ne sono derivate rappresentano non soltanto altrettante scoperte nella fisiologia degli Insetti, ma anche risultati di portata biologica generale. Un panorama è offerto dal recente volume edito da J. Beament e J. E. Treherne, Insects and physiology (1967), mentre parecchi articoli dell'ultimo decennio, pubblicati su numerose riviste e soprattutto su ‟Advances in Insect physiology", aggiornano sui principali problemi fisiologici e biochimici riguardanti gli Insetti.
Vediamo anzitutto quali sono le acquisizioni di base relative alla composizione chimica degli Insetti.
Per quanto riguarda i carboidrati (v. Wyatt, 1967), il glucosio, che pure è d'importanza fondamentale per il loro metabolismo, è tuttavia presente e dosabile in essi generalmente solo in quantità piuttosto limitate, in quanto lo zucchero tipico dell'emolinfa degli Insetti è il suo dimero trealosio; sono inoltre presenti, in relazione alla dieta, fruttosio, sucrosio, maltosio, allobiosio. Trealosio, glucosio e fruttosio sono gli zuccheri tipici di tutti i tessuti degli Insetti; il primo è presente durante l'intera vita, in quantità crescenti dalle uova alla pupa e decrescenti nell'adulto. Oli zuccheri, marcatamente il glucosio, vengono assorbiti dal mesointestino e soprattutto dai ciechi, passano nell'emolinfa e vengono convertiti in trealosio. Quest'ultimo, la cui concentrazione nel sangue è mantenuta costante, presenta un regolare metabolismo: viene infatti idrolizzato dall'enzima trealasi in due molecole di glucosio (trealosio + H2O → 2 glucosio) che poi è utilizzato come tale e, d'altro canto, può essere sintetizzato a partire dal glucosio, il quale, a sua volta, può avere origine dal tipico polisaccaride di riserva, che anche negli Insetti è il glicogeno, per azione della trealosio-isomerasi. Il glicogeno, accumulato soprattutto nel corpo grasso e nei muscoli, è sintetizzato a partire dal glucosio per azione della glucosiosintetasi, e può essere scisso liberando glucosio a opera sia della glicogenofosforilasi, sia della amilasi, presente negli Insetti, oltre che nell'intestino ove esplica le ovvie funzioni digestive, anche nell'emolinfa. Negli Insetti ibernanti il glicogeno viene convertito in sorbitolo e glicerolo, che, ostacolando il congelamento dell'emolinfa, determinano un aumento della resistenza al freddo. Un altro importantissimo carboidrato, presente nella cuticola in associazione con proteine per formare una glicoproteina, è la chitina o poli-acetilglucosammina, sintetizzata nell'epidermide dalla chitinsintetasi a partire da UDP-glucosio e UDP-acetilglucosammina presenti nell'emolinfa: la chitina, in quanto digerita da chitinasi e riassorbita non solo in occasione delle mute, ma anche per fornire glucidi durante i digiuni prolungati, deve essere considerata anche un polisaccaride di riserva. Un altro importante polisaccaride degli Insetti è l'acido ialuronico, presente nella membrana peritrofica che tappezza il lume mesointestinale e che è uno dei componenti della sostanza cementante dei connettivi.
I lipidi costituiscono un'altra importantissima categoria di sostanze contenute nell'organismo degli Insetti, rappresentandone in peso una percentuale che va da meno dell'1% (ape adulta) a oltre il 5o% (alcune larve di Lepidotteri): fra questi estremi esiste tutta una gamma di variabilità, ovviamente dipendente dallo stadio, dalla specie, dalle condizioni dietetiche (v. Gilbert, 1967). Un forte impulso allo studio qualitativo dei lipidi degli Insetti è stato dato dalla scoperta che gran parte degli ormoni che regolano le metamorfosi sono sostanze lipoidee.
La maggior componente lipidica dell'organismo degli Insetti è rappresentata da trigliceridi, che costituiscono una notevole sorgente di energia. Importanti sono anche gli acidi grassi liberi, la cui catena può essere costituita da 2 sino a 34 atomi di carbonio. I lipidi, introdotti nell'intestino come trigliceridi e digeriti da apposite lipasi in mono o digliceridi o in acidi grassi liberi, penetrano poi attraverso la parete intestinale nell'emolinfa, ove sono di solito coniugati con proteine; nel corpo grasso vengono quindi ricostituiti i trigliceridi, che, come si è detto, rappresentano una sorgente di energia, la cui liberazione si compie per l'idrolisi operata da lipasi extradigestive, generalmente a livello della trioleina o della tributirrina. Gli acidi grassi liberano energia per ossidazione: il processo viene utilizzato soprattutto nello sviluppo embrionale, essendo le uova ricchissime di lipidi: questi forniscono fino a 2/3 dell'energia necessaria all'embriogenesi e al metabolismo dei muscoli di volo. Oli acidi grassi possono venire sintetizzati negli Insetti secondo le vie metaboliche note per gli altri animali in genere e, a partire da essi, oltre ai trigliceridi, vengono sintetizzati i fosfolipidi, la cui importanza è in generale essenziale per il metabolismo cellulare.
Un'importante sede di concentrazione di lipidi è la cuticola, ove è presente uno strato di ‛cere' composto per il 75% da idrocarburi, e per il resto da acidi grassi, aldeidi alifatiche ed esteri: questo strato è responsabile della prevenzione dalla disidratazione dell'organismo degli Insetti. Idrocarburi e cere fanno parte di una significativa frazione dell'emolinfa; le cere inoltre sono secrete in abbondanza dalle femmine degli Imenotteri aculeati. Sembra che la sintesi delle cere avvenga in particolari cellule isolate, sottostanti all'epidermide, dette enociti, a partire da acetato ottenuto dal corpo grasso.
Un'importantissima categoria di sostanze lipoidee tipiche degli Insetti è quella degli isoprenoidi, che includono steroidi e terpeni. Molte di queste sostanze sono ormoni tipici degli Insetti. Poiché gli Insetti, a differenza dei Vertebrati, sono incapaci di convertire semplici precursori (per es., acetato) in steroli, questi devono venire assunti con la dieta da sostanze vegetali o animali; colesterolo, o altri steroli, vengono poi utilizzati come elementi per le strutture cellulari, o da particolari cellule come sostanze che immesse in circolo o addirittura liberate all'esterno funzionano come ormoni o feromoni. Le più note di queste sono l'ormone del cervello, un tempo ritenuto essenzialmente colesterolo, ma non ancora completamente definito dal punto di vista chimico; l'ormone della muta o ecdisone, steroide fortemente polare a 5 idrossili, derivato dal colesterolo; l'ormone giovanile, simile al farnesolo, ma con struttura chimica ancora sconosciuta; i feromoni sessuali, secreti dalle ghiandole addominali di tutti gli Insetti, e la ‛sostanza della regina' secreta dalle ghiandole mandibolari dell'ape regina: tutte sostanze lipoidee ora chimicamente note e anche artificialmente sintetizzabili (v. ormoni negli invertebrati).
Le proteine sono senza dubbio la categoria di sostanze più importanti per l'edificazione e il funzionamento del corpo degli Insetti: il metabolismo degli amminoacidi, recentemente riesaminato da P. Chen (v., 1966), sta infatti alla base di tutte le trasformazioni che l'insetto subisce, dalla vita embrionale allo stadio di adulto. Nell'uovo, al momento della fecondazione, sono presenti pochissimi amminoacidi liberi, che aumentano poi in numero e in quantità con il progredire dello sviluppo, attraverso le importanti tappe della gastrulazione, della formazione del mesenteron e di quella degli organi dorsali: questi amminoacidi derivano dall'idrolisi delle proteine del vitello, e servono all'edificazione sia delle principali proteine strutturali (l'artropodina del tegumento, il collagene dei tessuti di sostegno, la miosina, l'actina, la tropomiosina e la troponina dei muscoli ecc.), sia delle numerose proteine enzimatiche che pure aumentano in rapida progressione (le fosfatasi acide, le proteasi, gli enzimi respiratori ecc., cioè più o meno le stesse presenti negli altri animali). È importante ricordare che alcune di queste ultime, soprattutto gli enzimi digestivi quali proteasi, lipasi, ialuronidasi, entrano a far parte dei veleni degli Insetti e vengono secrete in quantità eccezionali in particolari momenti (v. Beard, 1963). Durante lo stadio larvale le esigenze dell'accrescimento determinano un'enorme richiesta di amminoacidi, utilizzati sia per la sintesi delle proteine, sia come elementi d'importanti vie metaboliche; alcuni amminoacidi, sono contrassegnati da un carattere d'indispensabilità strettamente specifica, mentre altri sono necessari per tutti gli Insetti. Tutti gli amminoacidi introdotti con la dieta si ritrovano liberi nell'emolinfa, ma il sistema escretore, ed essenzialmente i tubi malpighiani, praticano una depurazione che il retto ulteriormente seleziona riassorbendo alcuni elementi. Le più tumultuose sintesi proteiche avvengono nella fase che precede ciascuna muta, quando è sintetizzata l'artropodina dermascheletrica, e in quella fase che in molte larve precede la filatura del bozzolo, quando sono sintetizzate le proteine della seta. Durante l'impupamento la distruzione di molti tessuti larvali (istolisi) e la differenziazione delle strutture dell'adulto (istogenesi) danno luogo a drammatici cambiamenti sia nel corredo di amminoacidi liberi e proteine, sia nell'attività di particolari enzimi (tipica la curva a U della respirazione durante lo stadio pupale). L'adulto edifica nuove proteine per il mantenimento del proprio organismo e soprattutto sintetizza le proteine collegate con la riproduzione: quelle del vitello delle uova, quelle secrete dalle ghiandole accessorie degli organi genitali, ecc., con evidenti differenze, nello spettro di amminoacidi liberi nell'emolinfa, fra maschi e femmine.
La sintesi proteica è RNA dipendente. È stato anche dimostrato che l'attività di diversi ormoni che regolano la metamorfosi, la riproduzione o lo sviluppo dell'uovo si esplica mediante il controllo del metabolismo dell'RNA e quindi della sintesi delle proteine specifiche.
Questa è la via attraverso la quale si manifestano le mutazioni. Lo studio dei mutanti biochimici è uno dei campi d'avanguardia dell'entomologia attuale e ha consentito basilari acquisizioni di genetica molecolare.
Quasi tutte le proteine degli Insetti, dal collagene all'elastina, alle proteine muscolari, e perfino a quelle del vitello delle uova, sono simili a quelle degli altri animali. Un problema particolare è invece rappresentato dalle proteine tegumentali (artropodine e resiline) e da quelle della seta, ritenute specifiche degli Artropodi: entrambe le categorie meritano un discorso speciale. Col nome di artropodina G. Fraenkel e K. M. Rudall nel 1947 indicarono quel miscuglio di proteine che sono frammiste alla chitina nel tegumento degli Insetti. Si è ora accertato (v. Rudall, 1963) che questa miscela è in realtà più largamente diffusa nel regno animale e consta di cinque frazioni chimicamente definibili, ognuna con diversa costituzione amminoacidica; esse sono capaci di legarsi chimicamente alla chitina, costituendo così una glicoproteina, e possono venire stabilizzate e indurite mediante processi di tannazione chinonica. L'artropodina così tannizzata prende il nome di sclerotina. Pur se alcune sue qualità ricordano quelle della sericina e altre quelle del collagene, l'artropodina è nettamente diversa da queste due proteine e merita ulteriori studi. La resilina (v. Andersen e Weis-Fogh, 1964) è una proteina strutturale responsabile delle proprietà di alcune zone della cuticola altamente elastiche, quali legamenti, tendini ecc., chimicamente simile all'elastina, ma ricca di residui glicidici e priva di residui contenenti zolfo; il suo carattere fisico essenziale è, come per l'elastina, quello di una tessitura isotropa tridimensionale di catene polipeptidiche, tenute insieme da legami covalenti trasversali stabili. I suoi precursori sono delle proteine lineari solubili, non ancora isolate.
La seta è una stravagante mescolanza di sostanze diverse, in genere tutte ‛estramurali', utilizzate per la tessitura di bozzoli, nidi, filamenti di attacco. Secondo K. M. Rudall e W. Kenchington (v., 1971), l'insetto per filare può utilizzare svariate sostanze: di solito proteine, ma talora anche polisaccaridi o idrocarburi, disposti generalmente in fibre, ma anche in nastri o lamine. Una definizione chimica della seta è perciò impossibile; se ne può fornire solo una funzionale. Le proteine fibrose (fibroine) delle sete degli Insetti possono essere classificate in otto gruppi, ciascuno caratterizzato da una progressiva riduzione nei residui di glicina (da 2/3 a 1/10); di uno di essi, il terzo, fanno parte particolari sete, di tipo collagene, contenenti circa un terzo di glicina. In genere le catene proteiche della fibroma sono in forma β: i Tentredinidi, quando non hanno sete di tipo collagene, le hanno in forma α, e così le Mantidi, che con catene proteiche di questo tipo tessono le ooteche. Alcuni Coleotteri (Ptinus, Prionomerus) secernono probabilmente dall'intestino sete polisaccaridiche (di chitina); le larve delle Crisope elaborano, a livello dei tubi malpighiani, sete di tipo nylon, con condensazione di dicarbossili e di diamminoidrocarburi.
6. La fisiologia degli Insetti
Parallelamente al progredire delle conoscenze sulla morfologia funzionale e sulla biochimica degli Insetti, si registrava un progresso della fisiologia. Le più significative acquisizioni in campo entomologico, riguardanti i problemi inerenti alla particolarissima endocrinologia, alcuni peculiari aspetti del sistema nervoso, l'escrezione e con essa il controllo ionico e osmotico, hanno rappresentato validi contributi recati dallo studio degli Insetti alla soluzione di problemi generali e speciali. Per altri argomenti, invece, quali la contrazione muscolare, gli studi condotti sugli Insetti hanno semplicemente convalidato le conoscenze acquisite con le ricerche condotte su altri animali. Infine, per una particolare branca della fisiologia, l'etologia, si deve addirittura riconoscere negli Insetti il soggetto principale per l'impostazione sperimentale del problema.
Appena tratteggiato può essere qui il suggestivo quadro del controllo osmotico, ionico e idrico, e dell'escrezione negli Insetti, il cui studio iniziato da Wigglesworth negli anni trenta è stato di recente completato da numerosi autori (v. Shaw e Stobbart, 1963; v. Berridge, 1967). L'enorme adattabilità a tutti gli ambienti, anche ai più aridi e salmastri, di questi animali che tuttavia in moltissimi stadi preimmaginali vivono in acqua dolce, esige evidentemente potentissimi meccanismi di trasporto attivo di acqua e ioni attraverso gli epiteli. L'emolinfa che circola in tutto l'organismo è continuamente controllata dai tubi malpighiani e dagli altri organi escretori, e la perdita di acqua è costantemente limitata dall'impermeabilità del tegumento, dai sistemi di chiusura di alcuni suoi orifizi e dai sistemi di riassorbimento a invaginazioni tubulari submicroscopiche che fasciano tutti gli epiteli prossimi allo sbocco all'esterno (retto, dotto impari dei genitali, sbocchi di ghiandole, ecc.). Il tubo malpighiano, organo escretore completo, ma abbastanza semplice da manipolarsi, rispetto al convolutissimo rene dei Vertebrati, ha offerto un eccellente modello per lo studio citologico dell'escrezione, mediante l'associazione della microscopia elettronica con i sistemi di microanalisi: è stato così possibile da un lato collegare con il problema del controllo osmotico e ionico quello della escrezione azotata, che pure ha svelato negli Insetti una vastissima gamma di comportamenti, dal sistema uricotelico a quello uricolitico con, formazione di urea, a quello ammoniotelico (v. Bursell, 1967); e dall'altro, mettere in evidenza lo strettissimo collegamento tra caratteristiche dell'escrezione ed ecologia, individuabile in specie sistematicamente affini, ma ecologicamente diverse. Fra le numerose branche dell'entomologia fisiologica, lo studio dell'escrezione in senso lato è forse quello rimasto più aderente alla collaborazione con l'entomologia di campagna.
Come si è già detto, ancora poco si sa sulla costituzione chimica dei più importanti ormoni degli Insetti, tutti steroidi, mentre più conosciuta è la loro funzione, per la prima volta chiarita da V. B. Wigglesworth negli anni quaranta, e ora oggetto di numerosi scritti e rassegne, fra le quali magistrale quella del medesimo autore (v. Wigglesworth, 1964). Il sistema nervoso è il primo organo endocrino: la pars intercerebralis secerne l'ormone del cervello (quello colesterolo-simile) responsabile della stimolazione della ghiandola protoracica, la quale a sua volta secerne l'ecdisone o ormone della muta, il meglio noto chimicamente, che agisce sia sulle cellule epidermiche stimolandole a secernere una cuticola ‛adultoide', sia sulle gonadi e sull'intero organismo, e la cui assenza provoca diapausa, arresto di sviluppo sessuale ecc. Inoltre, la parte anteriore della catena ganglionare addominale elabora l'ormone diuretico scoperto da S. H. Maddrel nel 1962; l'ultimo ganglio addominale secerne l'ormone tannizzante, responsabile della sderificazione delle proteine, scoperto da R. R. Mills nel 1965, e il ganglio sottoesofageo secerne un ormone della diapausa (v. Hasegawa, 1952); l'epidermide è pure raggiunta da fibre nervose che trasportano direttamente un neurosecreto, così come avviene per i muscoli, la cui ciclica involuzione è pure controllata da un ormone. Tuttavia, le attività di questi ormoni rappresentano in generale dei problemi ancora insoluti. Tutto sommato, il meglio noto dal punto di vista funzionale (anche se chimicamente ancora non definito) è l'ormone giovanile o neotenina (il problema è stato riesaminato da V. J. A. Novak, v., 1967), secreto da due particolari organi neuroemali detti corpora allata, il quale contrasta l'ecdisone a livello dell'epidermide, stimola il mantenimento di caratteri giovanili, regolando direttamente o indirettamente proteosintesi, respirazione, mute, secrezione di seta e feromoni, comportamento ecc. Esso agisce come riattivatore di segmenti di DNA inattivati, al contrario dell'ecdisone che non è altro che un derepressore. I corpora allata, probabilmente sempre attraverso l'ormone giovanile o neotenina, regolano anche lo sviluppo delle gonadi, ma l'intricato complesso di queste interazioni abbisogna ancora di molto studio.
Il sistema nervoso degli Insetti, pur essendo uno dei più complessi ed evoluti tra quelli degli Invertebrati, possiede alcune caratteristiche che lo hanno reso assai utile per la soluzione di problemi di neurofisiologia generale (v. Treherne, 1967): esso è infatti metamerico, ed è perciò fornito di gangli relativamente piccoli e semplici, solidi, immersi in un'emolinfa che penetra nel loro interno diffondendosi attraverso i tessuti superficiali e nella quale sono presenti il sodio e il potassio che regolano le attività elettriche delle cellule nervose. È stato così possibile identificare nelle varie pareti che delimitano i gangli (cellule gliali, cellule perineurali, guaina collagene, guaina adiposa) le diverse attività trofiche (v. D. S. Smith, 1967), di filtro, addirittura di regolazione del medium extracellulare. Numerosi sono però i problemi tuttora insoluti nella neurologia entomologica (v. Edwards, 1967): non si conoscono le modalità dell'incremento delle fibre nervose durante l'accrescimento dei muscoli, marcatissimo nel periodo dello sviluppo larvale e particolarmente durante l'impupamento, quando compaiono addirittura nuove parti corporee; non si conoscono i meccanismi della regolazione dell'accrescimento del neurone e della costellazione di cellule gliali che lo circondano; s'ignora in quale modo dopo amputazione le fibre afferenti rigeneranti possano penetrare nel ganglio e quivi stabilire contatti sinaptici nella giusta posizione.
Lo studio dei raffinatissimi organi di senso degli Insetti ha aperto un vastissimo campo d'indagini, ove la ricerca è risultata più agevole per quanto riguarda l'enorme gamma di meccanocettori semplici dagli apparati esterni assai stravaganti, fatti di cuticola a sottilissima parete (v. Slifer, 1967), più complessa invece nel settore dei meccanocettori in batteria costituenti veri e propri ‛orecchi' (anche se dislocati in zone del corpo per noi inconsuete) in grado di assicurare agli Insetti un enorme vantaggio selettivo nella vita sulla terraferma, eguagliato solo dai Vertebrati (v. Jurato, 1967). La problematica appare particolarmente vivace e suggestiva nel campo dei fotocettori, che negli Insetti, come è noto, hanno raggiunto specializzazioni e acutezza favolose: per lungo tempo, infatti, non era stato possibile spiegare come gli innumerevoli occhi semplici (ommatidi), che costituiscono i famosi occhi composti tipici degli Artropodi, fossero in grado di comporre le singole immagini in un unico quadro. Sia negli Artropodi sia nei Vertebrati il fotocettore elementare presenta l'identica struttura a bastoncelli ordinati, il rabdoma, e il composto fotosensibile, il retinene, derivato dalla vitamina A, è il medesimo; le singole cellule retiniche di un Insetto, inoltre, reagiscono alla luce con minuti cambiamenti di potenziale come tutte le normali fotocettrici. La caratteristica del tutto particolare è rappresentata dal fatto che ogni ommatidio ha un proprio apparato diottrico, in genere cuticola trasformata, ed è separato dagli ommatidi vicini da strati di pigmento; la spiegazione più immediata di ciò fu avanzata da J. Müller nel 1840, il quale prospettò l'ipotesi della composizione di un'immagine mosaico a livello cerebrale. Le acquisizioni successive hanno consentito di chiarire il fenomeno (v. Burtt, 1967): intanto le cellule fotocettrici degli Insetti hanno una capacità di adattamento in passato impensabile, spostando dalla luce all'ombra pigmenti e mitocondri, addirittura variando la velocità nel cambiamento di potenziale, e agendo tutte in perfetta sincronia; pare poi che le singole cellule retiniche di ciascun ommatidio siano specializzate per un diverso colore e che lo stesso avvenga per l'analisi della luce polarizzata. Tuttavia, è confermato che il singolo ommatidio agisce come un'unità fisiologica indipendente. Una probabile ipotesi di valore generale ammette che una visione per sovrapposizione, nella quale gli ommatidi agiscono in gruppo ottenendo la possibilità di una più alta risoluzione, consenta l'identificazione preliminare degli oggetti lontani e che, dopo che l'insetto si è avvicinato, entri in giuoco la visione a mosaico, operata da altri fotocettori sempre nei medesimi ommatidi. L'integrazione dei due sistemi di visione, nonché quella dei dati ottici con altri dati sensoriali, avviene molto in superficie nel sistema nervoso: si pensi che unità miste visuali e acustiche sono state trovate da G. A. Horridge a livello della seconda regione sinaptica del lobo ottico. Ma molto poco sappiamo sulla trasmissione di tutti questi tipi d'impulsi alle altre parti del sistema nervoso. La problematica globale dell'integrazione dei diversi dati sensoriali e delle reazioni a essi conseguenti appartiene a una delle discipline più antiche dell'entomologia, da poco tornata in grande evidenza: l'etologia.
7. L'etologia entomologica
Il tentativo di descrivere, oltre la forma, anche le abitudini di un Insetto costituì uno dei primissimi approcci al problema entomologico generale. Come abbiamo visto, furono Redi e Vallisnieri, Aldrovandi e Réaumur a scoprire e a diffondere il gusto di quest'indagine. Indagine semplice, che richiede mezzi tecnici esigui, in grado di valorizzare l'osservazione sporadica, per molti secoli di difficile documentazione e quindi sfuggente a verifiche sicure; indagine che stimolò numerosi cultori trasportandoli spesso ai limiti della fantasia e della poesia: primeggiò fra tutti il francese J.-H. Fabre (1879-1907). Il complesso delle nozioni sui costumi degli Insetti ai primi decenni di questo secolo era enorme, con una grande quantità di fatti veri mescolati a fantasie (la cui verifica è ancora ben lontana): ne risultava quindi un quadro globale impreciso, d'interpretazione assai dubbia.
La nascita dell'etologia come disciplina rigorosamente sperimentale, su basi fisiologiche, è molto recente. Già nella prima edizione del trattato di fisiologia degli Insetti di V. B. Wigglesworth (v., 1939) compare un capitolo sul behavior, disciplina che dovrebbe condurre alla scomposizione di tutti i movimenti di un intero animale in una serie di riflessi. Il tentativo non è semplice, a causa della grande varietà delle terze componenti variabili che intervengono fra stimolo e risposta allo stimolo. Tuttavia, questa è la strada più volte indicata da J. S. Kennedy (v., 1967), il quale così testualmente si esprime: ‟Un'importante caratteristica dell'entomologia contemporanea è costituita dal fatto che lo studio del comportamento degli Insetti è divenuto una disciplina indipendente, con proprie acquisizioni scientifiche che da un lato sfiorano le vicine discipline della fisiologia e dall'altro l'ecologia, pura o applicata. È pur vero che, per lunga tradizione, esso ha anche un terzo legame, attraverso l'antropomorfismo, con la psicologia, il che continua a disturbare il contenuto fisiologico e quello ecologico; ma il comportamento ha già rapporti con la neurofisiologia, e viceversa, specialmente a livello di particolari, determinate risposte nelle quali può essere interpretato fisiologicamente, il che equivale ad aprire la porta all'analisi neurofisiologica di più complesse interazioni comportamentali. Gli studi sul behavior aiutano anche a cambiare il contenuto di alcuni concetti ecologici generali, quali l'adattamento, la migrazione e la competizione infraspecifica, ove i modelli meccanicisti finora esistenti cedono il passo ad altri, più raffinati, di biologia dell'adattamento. Nell'ecologia applicata, intesa come lotta contro le infestazioni, il comportamento è divenuto il punto di maggiore interesse e si può sperare che a questo riguardo le ulteriori raffinatezze non rimarranno così largamente ancorate alla chimica" (v. Kennedy, 1972, p. 337).
Fra i numerosi capitoli che l'etologia entomologica ha in questi ultimi anni approfondito, due appaiono di particolare interesse: l'orientamento (e la possibilità di comunicare informazioni relative a esso), e l'apprendimento (con la conseguente possibilità di memorizzare). Per orientamento, le cui conoscenze sono state di recente riassunte da L. Pardi (v., 1957), da H. Mittelstaedt (v., 1962) e da R. Jander (v., 1961), s'intende il controllo attivo delle relazioni spaziali da parte di un organismo: esso può essere ‛aperto' (cioè al di là dello stimolo ricevuto, come nel caso delle lucciole che si voltano lentamente verso un rapidissimo flash luminoso, completando l'operazione molto tempo dopo che lo stimolo è scomparso), o ‛chiuso' (cioè esattamente corrispondente allo stimolo ricevuto, che può essere mobile o fisso, in un'area limitata o circolare).; quest'ultimo senz'altro più complesso e comprendente diverse capacità. Anzitutto gli Insetti che si orientano verso una sorgente luminosa sono capaci di mantenere un angolo fisso fra asse del corpo e sorgente e, se lo sperimentatore fa variare questo angolo, essi lo ripristinano, girando su se stessi per la via più breve e cioè per angoli inferiori ai 180°; se la sorgente cambia di 180°, l'insetto ruota di un angolo piatto in una qualunque delle due direzioni. Generalmente le variazioni d'intensità dello stimolo luminoso non modificano la corsa dell'insetto; l'altezza della sorgente luminosa non esercita influenza sugli Insetti che si muovono su piani orizzontali, ma solo su quelli che si muovono su piani verticali. Quando sono forzati a passare rapidamente da orientazioni soltanto fototattiche su piani orizzontali a orientazioni soltanto geotattiche su piani verticali, gli Insetti stabiliscono identiche relazioni angolari a qualunque stimolo, luce o gravità. Inoltre, molte specie cambiano la loro corsa rispetto alla luce, compensando i movimenti dell'azimut solare, in modo da ottenere una direzione di navigazione costante; anche l'orientazione geotattica mostra un movimento giornaliero corrispondente e la luce polarizzata del cielo può essere usata come sostituto del sole. Api e formiche possono calcolare le loro coordinate in movimento e così tornare a casa dopo escursioni in territorio sconosciuto. Molte di queste conclusioni sono state ottenute da K. von Frisch, il quale ha pure dimostrato che le api sono in grado di comunicare alle compagne la direzione e la distanza di una sorgente di cibo mediante una particolare danza. Normalmente gli Insetti stabiliscono una pista semplicemente sprigionando feromoni ai quali i compagni di specie sono sensibili. Tutto questo significa possibilità di apprendimento.
I problemi relativi all'apprendimento e alla memoria negli Insetti sono stati discussi dal T. M. Alloway (v., 1972). Tutte le più importanti esperienze di addestramento sono state condotte su Blatte, Tenebrioni del grano, Drosofile e soprattutto formiche, vespe e api, e si è potuto dimostrare che tanto l'apprendimento quanto la memoria procedono per stadi successivi e che vi è un altissimo grado di specificità nei diversi stimoli che vengono fissati; sono stati individuati diversi fattori (temperatura, tempo) che giuocano sulla memorizzazione e si è anche dimostrato un immagazzinamento definitivo.
Ovviamente una forte componente di behavior è geneticamente acquisita, come lo è la facoltà di apprendere più o meno e memorizzare. Una recente discussione di A. W. Ewing e A. Manning (v., 1967) dimostra che ampie basi di comportamento non soltanto sono geneticamente fissate, ma rappresentano anche oggetto importante di selezione e di evoluzione e mutano ed evolvono in parallelo con la morfologia, che negli Insetti adatta rigidamente tutti gli stadi ad ambienti ben precisi. Il difficile e complesso studio sperimentale della genetica del comportamento ha condotto a inconfutabili acquisizioni, consentendo tra l'altro di dimostrare che sono geneticamente fissati in modo particolare tutti i comportamenti legati alla riproduzione, compreso il corteggiamento (tipico, il canto dei grilli), ciò che costituisce il presupposto fondamentale dell'isolamento riproduttivo come fattore di speciazione. Si comprende quindi come l'etologia debba essere considerata uno dei campi di maggiore attualità per lo studio della genetica e quindi dell'evoluzione, della filogenesi e perfino della sistematica (v. etologia).
8. Genetica e dinamica di popolazione
Nei primi decenni di questo secolo, dopo la riscoperta delle leggi di Mendel, lo studio sperimentale della genetica trovò negli Insetti un materiale di enorme importanza, che consentì fondamentali acquisizioni rendendo famose due scuole: quella di T. H. Morgan e quella, a noi contemporanea, di M. J. D. White. La fortuna degli Insetti come materiale per il genetista è legata a diversi fattori: essi sono facilmente allevabili, in poco spazio, con generazioni rapide e numerosa prole; sono contrassegnati da vari caratteri abbastanza ben definibili quantitativamente (numero delle faccette oculari, numero di peli e setole, ecc.); posseggono cromosomi abbastanza grandi, in basso numero; infine, le larve dei Ditteri sono caratterizzate da quei cromosomi giganti, politenici, che hanno consentito una verifica citologica della ricostruzione delle mappe cromosomiche dedotte dalla percentuale di crossing-over. Le fondamentali acquisizioni che i geni sono allineati sui cromosomi, che il cariotipo è stabile e anche, in gran parte, che il sesso è geneticamente determinato da precise coppie di cromosomi (non per nulla le tre formule maschio XY, maschio XO, maschio XX sono denominate rispettivamente tipo Lygaeus, tipo Protenor, tipo Abraxas) furono ottenute da ricerche eseguite su Insetti. A partire dagli anni quaranta il grosso sforzo della genetica si concentrerà sulla struttura chimica del materiale ereditario e sullo studio della sua mutabilità nelle grandi popolazioni, e gli Insetti non saranno più un materiale d'avanguardia. Tuttavia, la grande lezione di metodo impartita dalla scuola statunitense aveva convinto numerosi entomologi dell'importanza della genetica per l'entomologia, cosicché le ricerche continuarono su due filoni essenziali: studio al microscopio dei cariotipi, o citogenetica, come fondamento della sistematica e della scienza della riproduzione, con conseguenti acquisizioni sulla partenogenesi, sulla poliploidia, ecc.; e studio della dinamica delle popolazioni naturali con metodi matematici, con conseguenti interessi applicativi nella definizione delle epidemie, delle infestazioni e di più sottili fenomeni quali le naturali fiuttuazioni, che in molti Insetti presentano una base genetica.
La citogenetica è ormai strumento essenziale degli entomologi, come risulta da recentissime rassegne di M. J. D. White (v., 1957 e 1970), di S. O. Smith (v., 1960) e di molti altri autori. Per molte centinaia di specie il cariotipo è ormai noto e, grazie alle cognizioni relative al numero dei cromosomi, alla loro forma (dimensioni, posizione del centromero, costrizioni secondarie), alla forma dei nucleoli, alla quantità di DNA istofotometricamente determinata, è possibile una verifica della sistematica secondo due ordini di idee: filogenesi dei grandi gruppi e discriminazione di specie vicine altrimenti indistinguibili.
La dinamica delle popolazioni degli Insetti ha rappresentato un argomento principe per gli entomologi puri e applicati del dopoguerra. In una recente rassegna, H. O. Andrewartha e L. C. Birch (v., 1960) prendono in esame dettagliato le quattro componenti che costituiscono l'ambiente di ogni animale: clima, cibo, altri organismi e spazio. Il clima ha evidentemente un'importanza fondamentale, in quanto in genere gli Insetti sono organismi altamente specializzati per nicchie ecologiche assai precise; esso regola perciò le fluttuazioni numeriche di ciascuna specie, sia nei lunghi periodi geologici, con cambiamenti climatici relativamente stabili, sia in breve arco di tempo, durante un medesimo anno, magari in una sola generazione (v. Richards, 1961). Il cibo è un fattore limitante essenziale, data la notevole specializzazione anche nei suoi riguardi della generalità degli Insetti; in alcuni casi la spiccata monofagia ha consentito all'uomo di eliminare particolari specie dannose mantenendo le fonti di cibo molto sparse su vaste aree e sempre al di sotto delle soglie di sufficienza, come è stato per es. sperimentato contro la Glossina con il mantenimento di mandrie bovine in formazione sparse su enormi territori, anziché concentrate in grossi branchi. Altri organismi possono influenzare le popolazioni di Insetti in due modi: o usando il medesimo cibo (risultando cioè competitive), o interferendo direttamente con la specie (risultando cioè suoi parassiti o predatori). Nella vastissima gamma di possibilità, documentata dalle numerose ricerche iniziate già da lungo tempo, di particolare interesse appare il caso d'individui appartenenti alla medesima specie, che, oltre alla ovvia competitività al di sopra di un certo numero, possono anche esprimere un'aggressività particolare se opportunamente manipolati dall'uomo, come si verifica per es. quando esemplari irradiati o comunque geneticamente danneggiati siano immessi in popolazioni naturali allo scopo di deprimerne il livello. Lo spazio vitale, quarto fattore, agisce essenzialmente condizionando i precedenti due. La plasticità genetica delle specie che si muovono entro questi quattro fattori ambientali consente un certo ambito di variabilità in risposta alle variazioni delle componenti climatiche. Il genetista di popolazione studia appunto queste risposte e se ne vale allo scopo di prevedere infestazioni e di tentare di regolarne il numero. Attualmente, quindi, la genetica ecologica costituisce la più valida entomologia agraria o medica, tanto che si può affermare che interi complessi fenomeni, come quello del gregarismo delle locuste (B. P. Uvarov e sua scuola), sono stati risolti proprio su questa base (v. ecologia).
9. L'entomologia economica
a) Gli Insetti merceologicamente utili
Nella vecchia farmacopea erano elencati Insetti medicamentosi, tra i quali nel ruolo di protagonisti figuravano regolarmente le Cantaridi e i produttori di lacche alimentari (i Chermes); così pure, per lungo tempo, le cocciniglie sono state utilizzate dai tintori. Nell'epoca moderna, invece, gli Insetti ancora impiegati dall'uomo a scopo industriale sono praticamente soltanto il baco da seta e l'ape. T. Yokoyama (v., 1963) elenca diverse specie di bachi da seta: il Bombyx mori e i meno noti Antheraea pernyi, Antheraea yamamai, Phylosamia cynthia, Antheraea mylitta, fra le quali però soltanto la prima è stata industrialmente allevata e studiata. Già nel primo trattato sull'argomento, opera di un ignoto quattrocentesco italiano (v. Baccetti, 1965), apparivano delineate le problematiche basi della sericoltura: coltivazione dei gelsi, eterogeneità delle razze di bachi (se ne annoverano oggi un migliaio, il cui bozzolo differisce per i più vari caratteri), velocità di sviluppo del baco in rapporto alla quantità di alimento, qualità della seta in rapporto alla razza e al nutrimento, malattie del baco. Quest'ultimo capitolo è il più importante e ad esso la scuola italiana ha per il passato contribuito in modo determinante. È comunque superfluo osservare che attualmente l'importanza economica della sericoltura è assai diminuita. Intramontabile è invece quella dell'ape, animale dalle abitudini complicatissime per le peculiari caratteristiche della regolazione dei sessi mediante partenogenesi arrenotoca, facoltativa, della divisione in caste sulla base dell'alimentazione larvale, dell'elevatissima socialità con divisione del lavoro, del controllo del numero delle femmine riproduttrici e quindi della generale popolazione della società. Gli studi compiuti sotto l'impulso dell'importanza economica dell'ape hanno veramente consentito di gettare le basi della conoscenza dell'etologia degli Insetti. Anche nello studio delle api (v. Butler, 1956) importante è stato operare la discriminazione in razze, e individuare i caratteri del loro prodotto (miele e cera) in rapporto ai fiori bottinati; è così apparso sempre più evidente un legame fra pascoli e miele. Grandi progressi sono stati anche compiuti nelle conoscenze delle malattie dell'ape, che come tutti gli altri animali sociali è caratterizzata da un elevato grado di suscettibilità a gravissime virosi.
b) Insetti e agricoltura
È questo un problema antichissimo. Gli Insetti, infatti, hanno da sempre sottratto all'uomo enormi quantità di derrate alimentari e sono diventati progressivamente più dannosi man mano che l'uomo ha intensificato la propria agricoltura, concentrando piante della medesima specie, trasportando semi e piantine in lungo e in largo per tutto il mondo, immagazzinando derrate dei più vari tipi, eliminando specie competitrici, costruendo serre e ripari, assicurando in tal modo, senza pericoli e senza soste, cibo per gli Insetti. Così l'entomologia agraria ha assunto un'importanza sempre maggiore e ha richiesto, ovunque, a partire dagli anni trenta di questo secolo, sforzi addirittura immani. Gli Insetti possono danneggiare le piante in molti modi: i masticatori riducono il fogliame, scavano i fusti, distruggono i germogli e i fiori o addirittura i semi con conseguente limitazione della riproduzione, riducono le radici diminuendo in tal modo le capacità di assorbimento, rimuovono le riserve dai tuberi ostacolando così le possibilità di accrescimento, infine facilitano l'insorgenza di malattie fungine e batteriche; i succhiatori anzitutto diminuiscono il vigore delle piante riducendone la linfa, trasmettono virus e iniettano tossine, consentono la penetrazione di Funghi e Batteri e danneggiano inoltre foglie, fiori e frutti, riducendo anche la produzione di semi. Tuttavia, l'attività distruttiva degli Insetti sarebbe in equilibrio con le capacità di riproduzione e sopravvivenza delle piante, senza prevalenza dell'una rispetto alle altre, se l'uomo non avesse sempre, mai però come in questi ultimi decenni, profondamente alterato gli equilibri biologici di tutte le zone del pianeta. Così, quella che dovrebbe essere la normale presenza di Insetti su una coltura assume spesso il carattere di ‛infestazione', raggiungendo un livello massimo, economicamente insopportabile, tale da giustificare una spesa per la sua riduzione. In ciò consiste dunque il problema: se sia preferibile intervenire, e in tal caso quando e come, o piuttosto prevenire mediante un'accorta pianificazione delle colture fondata sulla previsione della dinamica delle popolazioni di Insetti, e sempre sopportando un certo grado d'infestazione. Comunque, il problema della lotta contro gli Insetti agrari non è dissimile da quello della lotta contro altre categorie di Insetti dannosi e verrà discusso globalmente in seguito (v. sotto, cap. 10).
c) Insetti e foreste
I danni che gli Insetti arrecano alle foreste consistono in decremento del patrimonio boschivo, degradazione della qualità del legno e deterioramento del legname tagliato (v. Oraham, 1956). I primi due tipi di danni possono essere determinati da qualunque Insetto, il terzo, ovviamente, solo da Insetti masticatori capaci di scavare gallerie. La problematica dell'entomologia forestale è naturalmente alquanto diversa da quella dell'entomologia agraria, in quanto ha per oggetto la difesa di un prodotto in linea di massima più povero, per il quale cioè più difficilmente si supera la soglia della convenienza economica di un trattamento a base di insetticidi. La pratica migliore è generalmente quella (v. Francke-Grosmann, 1963) di una gestione delle foreste intesa a prevenire il pullulare d'Insetti dannosi, che si attua regolando la qualità delle essenze, curando lo stato di salute delle piante e prestando molta attenzione ai nuovi alberi importati, che possono provocare effetti catastrofici se carichi d'Insetti nocivi per i quali non esistano in loco nemici naturali. In determinati casi è possibile addirittura introdurre nelle foreste alcuni nemici degli Insetti dannosi (Funghi o altri Insetti o Uccelli) e favorirne le condizioni di vita, sempre però esercitando un rigoroso controllo perché sia mantenuto il necessario equilibrio biologico, al fine di evitare la completa distruzione dei fitofagi che avrebbe come conseguenza la morte per inedia dei loro nemici. Raramente gli insetticidi sono stati adoperati in foresta e si può quindi affermare che, sotto molti aspetti, il selvicultore è stato uno fra gli entomologi agrari più illuminati.
d) Insetti e suolo
È questa la parte più nuova dell'entomologia applicata, in quanto l'enorme numero di Insetti, generalmente minuscoli, che soltanto in uno stadio o per tutta la vita si trovano nei diversi strati del suolo, è oggetto di un preciso studio sistematico da pochi anni. In un primo momento s'intendevano come Insetti del suolo agrariamente interessanti soltanto quelli che danneggiavano in qualche modo le radici delle piante, contro i quali furono escogitati insetticidi di vario tipo, il cui uso ebbe come risultato di alterare in modo spesso definitivo la microfauna presente nel medesimo suolo e ivi costituente uno dei punti base delle catene alimentari.
Fra gli Insetti, importantissimi abitatori dei suoli sono diversi gruppi di Atterigoti e numerose larve olometaboliche il cui effetto è generalmente più positivo che negativo; attualmente i programmi di difesa del suolo tendono a tenere nel massimo conto l'attività, ancora mal nota, dei piccoli Artropodi secondo linee di studio che ricalcano le ben note ricerche condotte sui lombrichi, i quali fin dai tempi di Darwin furono ritenuti i più utili custodi del suolo agrario.
Evidentemente la gestione del suolo, implicando il rispetto degli Artropodi utili e il controllo, possibilmente non chimico, di quelli dannosi, è un problema che esige una profonda conoscenza biologica ed ecologica di ciascun protagonista, anche perché il tipo di popolazione presente in un suolo può essere ritenuto un buon indice delle condizioni di salute del suolo medesimo. Il riconoscimento sistematico delle migliaia di minuscole specie di Insetti che vivono in un pugno di terra è opera della quale solo pochi specialisti sono capaci, e rappresenta uno dei tanti argomenti che in questi ultimi anni hanno nuovamente richiesto l'intervento dei vecchi sistematici dell'entomologia.
e) Insetti e virus
Si è già avuto modo di osservare che gli Insetti sono capaci di trasferire virus nocivi da una pianta all'altra, arrecando in questa maniera danni di notevole entità. Oltre che alle piante, gli Insetti possono anche risultare dannosi all'uomo, ed ai Vertebrati in genere, trasferendo virus responsabili delle più svariate malattie. La virologia degli Insetti è uno dei capitoli più nuovi nell'entomologia, perché soltanto alla fine dell'Ottocento, in coincidenza con la scoperta del famoso mosaico del tabacco, le malattie da virus furono riconosciute come una precisa categoria di disordini delle piante. La storia di un virus da quando viene assunto dall'insetto a quando viene inoculato in un altro ospite, dopo aver circolato nell'organismo dell'insetto stesso o essere semplicemente rimasto appeso alla sua superficie esterna, è un complicato compito della biologia, chiarito soltanto grazie all'impiego della microscopia elettronica e delle tecniche di coltura di cellule in vitro. In queste linee di ricerca emergono due scuole contemporanee: quella inglese di K. M. Smith e quella americana di K. Maramorosh. I virus possono penetrare negli Insetti nei modi più vari, sia venendo semplicemente assunti con l'apparato boccale (e questo è soprattutto il caso degli Insetti succhiatori), sia addirittura perché sono congenitamente presenti nell'uovo infettato nell'organismo materno; essi possono vivere nei nuclei, nei citoplasmi e persino in ammassi di cellule, e possono risultare dannosi o anche utili all'insetto stesso. Ma quelli che maggiormente interessano l'entomologia agraria sono i virus per i quali l'insetto rappresenta soltanto un vettore in grado di trasportarli da una pianta all'altra e da un animale all'altro.
Questi virus possono rimanere ancorati agli scleriti dell'apparato boccale o dell'ovopositore, oppure possono venire ospitati nelle ghiandole salivari o nell'intestino, giungendo addirittura a circolare nell'emolinfa. In alcuni casi la permanenza nell'organismo di un insetto è indispensabile al virus per una migliore propagazione e talora anche l'insetto vettore, oltre alla pianta, denuncia cambiamenti metabolici sensibili. Ma non tutti i virus sono dannosi all'agricoltura; numerosi virus poliedrici contenuti in vari vettori vengono usati per combattere pericolosi Insetti, e la virologia applicata (v. K. S. Smith, 1967) costituisce ora una delle problematiche entomologiche in via di maggiore sviluppo.
f) Insetti e Vertebrati
Si può dire che il problema dei danni diretti che gli Insetti sono in grado di arrecargli si è prospettato all'uomo fin dalle sue origini, avendo egli ereditato dai propri antenati scimmieschi un ricco corredo di pulci, pidocchi, cimici, mosche e zanzare che si sono man mano specializzati nei nuovi e sempre più confortevoli habitat umani; le stesse considerazioni valgono anche per quei Vertebrati che l'uomo ha addomesticato, alloggiandoli nelle stalle o accogliendoli addirittura nelle sue abitazioni.
Si usa distinguere gli Insetti dannosi per i Vertebrati in parassiti esterni e interni: fra i primi sono compresi tutti i succhiatori di sangue (Anopluri, Afanitteri, molti Emitteri, molti Ditteri) e i rosicchiatori di peli e penne (Mallofagi); dei secondi fa parte tutta la schiera dei Ditteri Estridi e Gastrofilidi, che passano gran parte della vita larvale entro il corpo di Mammiferi di grossa taglia. Ma l'aspetto di gran lunga più interessante del problema è senz'altro costituito dalla capacità di alcuni Insetti di trasmettere determinate malattie: esempi classici di ciò sono il plasmodio della malaria trasmesso dalla zanzara anofele, i Tripanosomi trasmessi dalla Glossina, la tenia del cane trasmessa dalla pulce. L'individuazione e la descrizione dei cicli di questi animali, ad alcuni dei quali sono legati i nomi di grandi parassitologi italiani come O. B. Grassi, furono importantissime conquiste della parassitologia tra la fine dell'Ottocento e l'inizio del Novecento e offrirono la possibilità di colonizzare aree prima impraticabili. Il nuovo capitolo che si sta aprendo, quello della trasmissione dei virus, processo nel quale è noto che sono implicati gli Insetti, anche se le idee, sia sulla specificità dei trasmettitori sia sui cicli, sono in generale alquanto confuse, è certamente uno dei più importanti della storia dell'entomologia e determinerà senza dubbio un forte rilancio dell'entomologia medica.
10. Il problema della lotta contro gli Insetti
Quando un intervento contro un determinato insetto dannoso appare economicamente sostenibile e risulta necessario occorre decidere cosa fare; sinora innumerevoli volte agricoltori e allevatori hanno dovuto prendere decisioni di questo tipo e le misure adottate hanno variato nel corso dei tempi.
Nel secolo scorso il problema appariva relativamente semplice. I pionieri dell'entomologia agraria (fra i quali un posto d'onore occupa il fiorentino A. Targioni Tozzetti, fondatore della Stazione di Entomologia Agraria del Ministero dell'Agricoltura) ricorrevano ai veleni più noti della farmacopea, somministrando agli Insetti in vari modi arseniati, arseniti, fluoruri, fosfuri, solfuri e anche nicotina, quassina, rotenone, curando tutt'al più la distinzione fra azione di contatto e d'ingestione e in quest'ultimo caso valutando la solubilità del prodotto, onde poterne favorire la penetrazione anche nell'interno degli apparati boccali succhiatori. A quell'epoca quindi la strategia della lotta era assai rudimentale e si cercava di annientare il maggior numero possibile di Insetti; d'altra parte, un controllo veniva effettivamente praticato soltanto in poche aree e non si ponevano grandi problemi di turbamento di equilibri biologici. È dagli inizi di questo secolo che i sempre crescenti traffici e spostamenti di piante vive o morte e di derrate da un capo all'altro del mondo hanno determinato lo spostamento di Insetti al di fuori dei luoghi di origine, ove erano in equilibrio con tutte le cause loro avverse che da sempre ne avevano condizionato l'evoluzione. In queste nuove condizioni il fitofago emigrante, di fronte al differente regime concorrenziale, o si trova male e soccombe, o si trova bene e, privo di parassiti specifici (che difficilmente seguono le vittime nelle emigrazioni), pullula. Questo secolo è stato appunto caratterizzato dalla comparsa in molte zone di numerosissimi nuovi Insetti dannosi, provenienti da regioni lontane dove erano quasi indifferenti. Alcuni entomologi illuminati (fra i quali va ricordato il già nominato A. Berlese) hanno propagandato un facile sistema di controllo consistente nello studio e nell'importazione di quei parassiti che nel suo stesso luogo d'origine limitano la diffusione dell'insetto espatriato (v. lotta biologica). Così, con intuizione eccezionale Berlese ne importò due che s'imposero subito, Prospaltella berlesei contro Diaspis pentagona (cocciniglia del gelso) e Novius cardinalis contro Pericerya purchasi (cocciniglia degli agrumi); un altro eccellente parassita fu importato in Francia da P. Marchai, Aphelinus mali, contro Eriosoma lanigerum, afide del melo. Ma la lista dei parassiti utilmente introdotti praticamente finisce qui: per tutta la prima metà di questo secolo, numerosissimi entomologi, anche i più famosi, hanno spostato qua e là i più svariati parassiti (spesso anche, goffamente, contro Insetti indigeni), ma senza conseguire risultati apprezzabili. Un risultato enorme, di diverso tipo, fu raggiunto da G. B. Grassi, che con la sua scuola sconfisse in Italia la fillossera della vite: dopo averne studiato completamente il ciclo biologico e aver scoperto la differente resistenza all'infestazione delle viti americane rispetto a quelle europee, caratterizzate dalla costante integrità rispettivamente delle radici e della parte aerea, egli prescrisse la coltura di vite europea su piede americano e, sostituendo gradatamente tutti i vitigni italiani, giunse a debellare un nemico dannosissimo. In questo modo venne dimostrata la possibilità per l'agricoltura di scegliere piante resistenti agli Insetti e si confermò la necessità di concentrare sempre più l'attenzione su un tipo di lotta condotta con mezzi naturali. Tuttavia, la pazienza di studiare le modalità reali ed efficaci di questa lotta fece difetto e d'altra parte durante la seconda guerra mondiale le industrie delle principali nazioni sintetizzarono, a volte senza scopi precisi, veleni di nuovo genere: conseguentemente, gli entomologi agrari ebbero a disposizione un'enorme gamma di sostanze nuove dalle proprietà insetticide elevatissime, comprendenti tutta la schiera dei cloroderivati (il cui capostipite è il DDT) e quella degli esteri fosforici, che irrorate largamente anche con l'impiego di aereoplani ed elicotteri (altri prodotti dell'industria bellica utilizzati in entomologia agraria negli anni quaranta) dettero al mondo l'illusione di una prossima vittoria facile e definitiva sugli Insetti. Ma nei decenni successivi si resero drammaticamente evidenti i troppo numerosi problemi che erano stati inizialmente trascurati. Anzitutto, non sarebbe stato possibile valutare fino a qual grado fosse nocivo l'accumulo nei tessuti umani di questi prodotti, che fanno ormai parte della dieta quotidiana, se non fosse trascorso un sufficiente periodo di tempo: soltanto negli ultimi anni sono emersi dati allarmanti sull'uso del DDT, cioè dopo almeno trent'anni di lento assorbimento, mentre per gli esteri fosforici, posti in commercio in un secondo momento, mancano ancora precise conoscenze ma non previsioni pienamente giustificate sulla loro tossicità. In secondo luogo è ben presto divenuto chiaro che le distribuzioni incontrollate di insetticidi non solo uccidono anche i parassiti degli Insetti nocivi, favorendo così l'esagerata e pericolosa proliferazione di altri Artropodi prima insignificanti perché naturalmente tenuti a freno, ma determinano anche la comparsa di razze resistenti (zanzare e mosche si attrezzano benissimo contro il DDT). Infine, è risultato evidente che se una specie si porta al di sotto di una certa soglia si affamano e si distruggono tutti i suoi parassiti, tanto che occorre poi tenerla sempre e necessariamente a freno con mezzi artificiali di lotta. Sempre più numerose furono le voci che si levarono negli anni cinquanta contro questo diluvio di insetticidi: in Italia una delle prime, la più intelligente e autorevole; fu quella di O. Grandi, entomologo dell'Università di Bologna. Ma la propaganda dell'industria, ormai lanciata sul redditizio mercato, ebbe ragione per molto tempo dei solitari predicatori del ritorno alla natura. Quando l'equilibrio è stato profondamente alterato e l'ambiente irrimediabilmente inquinato, ci si è finalmente accorti che era giunta l'ora di correre ai ripari e l'entomologia agraria ha incominciato a considerare l'entità di tutto il proprio affannoso lavoro e il valore delle proprie esperienze, felici e amare.
11. Il rilancio dell'entomologia
L'entomologia era negli anni cinquanta una disciplina vecchia, frammentata e superata come abitualmente accade alle scienze madri, dalle quali si distaccano continuamente nuovi rami mentre il vecchio tronco rimane sempre più nudo e insignificante: dapprima se ne distaccarono i genetisti, poi i fisiologi-biochimici, poi, con l'avvento della microscopia elettronica, perfino i citologi, infine addirittura gli entomologi applicati, che, in considerazione del vasto raggio di azione dei nuovi insetticidi, ritennero di poter fare a meno dei sistematici, di prescindere cioè da un preciso riconoscimento delle specie. Inoltre, alla sistematica smisero di affluire i giovani migliori, diretti verso altre frontiere della biologia. È chiaro dunque che non era più possibile parlare di un'entomologia come scienza unitaria. Il processo di ricostruzione si è realizzato nel corso degli anni sessanta e ha avuto il suo coronamento nel recente congresso mondiale tenutosi à Canberra nel 1972: è stata l'entomologia applicata e cioè la necessità economica di un controllo efficace, soprattutto in quelle regioni tropicali che l'uomo ha in questi anni deciso di abitare e non soltanto di sfruttare, che ha riunito le sparse membra della vecchia scienza. Si è così deciso che le infestazioni devono essere ‛gestite' e ‛previste' e non cancellate ciecamente; si è accertato che la fauna del suolo è basilare e che un insetto dannoso va considerato come il punto centrale di un enorme equilibrio biologico che vi ruota intorno; si è visto che ogni essere fa parte di una catena alimentare che non deve essere sconvolta. Tutto ciò ha portato a concludere che anzitutto occorre previamente elaborare un preciso catasto di ogni condizione, la cui indispensabile premessa è ovviamente la rivalorizzazione della sistematica in grande stile, e che i mezzi di lotta devono essere il più possibile specifici, in netto contrasto con la superficiale impostazione del dopoguerra. Conseguentemente si sono imposti all'attenzione nuovi mezzi: l'uso dei già ricordati ormoni regolatori della metamorfosi, dei quali recentemente sono state dimostrate le proprietà insetticide (v. Landa, 1972), che ha d'altra parte favorito il ritorno all'entomologia dei fisiologi; l'introduzione, nelle popolazioni di Insetti, di mutanti letali o sterili, naturalmente prodottisi o meglio ottenuti mediante irraggiamento o sterilizzazione chimica (da cui il famoso sistema dei maschi allevati e irradiati in laboratorio e poi liberati in eserciti, con il quale lo statunitense E. F. Knipling ha sradicato dal Texas un terribile tafano, la Callitroga hominivorax), che ha a sua volta richiamato citologi e genetisti. Una lotta di questo tipo, complicatissima, che tiene conto di tutti i fattori cercando di equilibrarli, è definita lotta integrata; può essere praticata soltanto con la collaborazione di entomologi di tutte le estrazioni che insieme si uniscano e gestiscano la popolazione di Insetti di territori il più possibile ampi. Lungo queste linee, che esigono ovviamente un'enorme organizzazione internazionale a tutti i livelli, si avvia l'entomologia degli anni settanta, mentre il lavoro improvvisato o isolato va rapidamente declinando.
12. Le nuove aperture della sistematica
A questo punto il discorso si chiude come si è aperto, con la perenne necessità di un catasto di specie quanto mai preciso, che tenga conto di tutti i caratteri che queste manifestano. La sistematica, a livelli assai alti, torna di moda e diventa indispensabile per tutte le operazioni di messa a punto ecologica e di stima di popolazione. Si esigono concetti il più possibile precisi, diagnosi nette, rapide, spesso da formulare su enormi masse di materiale: i metodi della sistematica vanno cioè cambiando. Un tempo era della massima importanza il colpo d'occhio dello specialista, che aveva in mente la fisionomia delle centinaia di specie componenti il gruppo da lui dominato, e che, con l'aiuto di collezioni di controllo, incasellava gli esemplari che via via incontrava in questa o in quella categoria; ne conseguiva che spesso, alla morte di un sistematico, tutto il lavoro doveva essere ricominciato da capo, con la creazione di una nuova memoria specializzata, dato che i libri, necessariamente impostati su un sistema di dicotomia, utilizzano un carattere alla volta e risultano sempre di gran lunga meno completi rispetto a quella massa d'informazioni sulle differenze fra determinate specie che il sistematico, spesso inconsciamente, utilizza tutte nel medesimo tempo. La sistematica numerica, di recente comparsa e di ancor più recente utilizzazione, colloca le specie non su un piano unico (secondo gli schemi della vecchia dicotomia), ma in uno spazio pluridimensionale, così che gli elementi della differenziazione risultano precisamente individuati (v. Sokal, 1967; v. Cole, 1969; v. Moss e Hendrickson, 1973). Risulta così chiaramente che nella complicata rete che ne emerge ben difficilmente ci si potrebbe orientare se non esistessero i calcolatori, i quali divengono sempre più indispensabili. In entomologia abbiamo già chiavi analitiche emergenti da calcolatori (v. Britton, 1972), il cui impiego risulta così prezioso, da un lato per elevare il livello di probabilità della diagnosi, dall'altro per facilitarla e abbreviarla (v. Wilkinson, 1972; v. Arnett, 1972) grazie alle possibilità di sfruttare in parallelo tutti i dati che la specializzazione della scienza attuale accumula in diversi settori. L'entomologia affronta dunque il suo quinto secolo di vita come scienza unitaria ed economicamente indispensabile.
bibliografia
Alloway, T. M., Learning and memory in Insects, in ‟Annual review of entomology", 1972, XVII, pp. 43-56.
Andersen, S. O., Weis-Fogh, T., Resilin, a rubberlike protein in Arthropod cuticle, in ‟Advances in Insect physiology", 1964, II, pp. 1-65.
Andrewartha, H. G., Birch, L. C., Some recent contributions to the study of the distribution and abundance of Insects, in ‟Annual review of entomology", 1960, V, pp. 219-242.
Arnett, R. T., Entomological information storage and retrieval, particularly for systematic entomology, in 14th International Congress of Entomology. Abstracts, Canberra 1972, p. 90.
Baccetti, B., Lo stroma di sostegno degli Insetti esaminato a luce polarizzata, in ‟Redia", 1956, XLI, pp. 259-276.
Baccetti, B., Trattatello di apicoltura, del porre i mori e del porre i bigatti, manoscritto inedito quattrocentesco, in Atti dell'Accademia Nazionale Italiana di Entomologia. Memorie, Bologna 1965, pp. 5-34.
Baccetti, B., Gli Insetti visti al microscopio elettronico, in Atti dell'Accademia Nazionale Italiana di Entomologia. Rendiconti, Bologna 1996, XIV, pp. 17-43.
Baccetti, B., Capra F., La microscopia elettronica a scansione nella sistematica entomologica, in Atti dell'Accademia Nazionale Italiana di Entomologia. Rendiconti, Bologna 1970, XVIII, pp. 1-16.
Beard, R. L., Insect toxins and venoms, in ‟Annual review of entomology", 1963, VIII, pp. 1-18.
Berlese, A., Gli Insetti, vol. I, Milano 1909, vol. II, Milano 1925.
Bernard, C., Introduction à l'étude de la médicine expérimentale, Paris 1865.
Berridge, M. J., Ion and water transport across epithelia, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 329-347.
Britton, E. B., The use of computer-generated keys, in 14th International Congress of Entomology. Abstracts, Canberra 1972, p. 91.
Bursell, E., The excretion of nitrogen in Insects, in ‟Advances in Insect physiology", 1967, IV, pp. 33-67.
Burtt, E. T., Some problems of Insect vision, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 217-232.
Butler, C., Some recent advances in apicultural research, in ‟Annual review of entomology", 1956, I, pp. 281-298.
Chen, P. S., Amino acid and protein metabolism in Insect development, in ‟Advances in Insect physiology", 1966, III, pp. 53-132.
Cole, A. J. (a cura di), Numerical taxonomy, London-New York 1969.
Edwards, G. A., Insect micromorphology, in ‟Annual review of entomology", 1960, V, pp. 17-34.
Edwards, J. S., Some questions for the Insect nervous system, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 163-173.
Ewing, A. W., Manning, A., The evolution and genetics of Insect behaviour, in ‟Annual review of entomology", 1967, XII, pp. 471-494.
Fabre, J.-H., Souvenirs entomologiques, 10 voll., Paris 1879-1907.
Fraenkel, G., Rudall, K. M., The structure of Insect cuticles, in ‟Proceedings of the Royal Society" (Series B), 1947, CXXXIV, pp. 111-143.
Francke-Grossmann, H., Some new aspects in forest entomology, in ‟Annual review of entomology", 1963, VIII, pp. 415-438.
Frisch, K. von, Lindauer, M., The ‛language' and orientation of the Honey bee, in ‟Annual review of entomology", 1956, I, pp. 45-58.
Gilbert, L. I., Lipid metabolism and function in Insects, in ‟Advances in Insect physiology", 1967, IV, pp. 69-211.
Gilby, A. R., Lipids and their metabolism in Insects, in ‟Annual review of entomology", 1965, X, pp. 141-160.
Goidanich, A., Entomologia, in Enciclopedia agraria italiana, vol. III, Roma 1957, pp. 856-863.
Graham, S. A., Ecology of forest Insects, in ‟Annual review of entomology", 1956, I, pp. 261-280.
Grandi, G., Introduzione allo studio dell'entomologia, 2 voll., Bologna 1951.
Grandi, G., Istituzioni di entomologia generale, Bologna 1966.
Grassi, G. B., Contributo alla conoscenza delle Filosserine ed in particolare della Fillossera della vite, Roma 1912.
Hasegawa, K., Studies on the voltinism in the silkworm Bombyx mori L. with special reference to the organs concerning determination of voltinism, in ‟Journal of the Faculty of Agriculture of the Tottori University", 1952, I, pp. 84-124.
Henning, W., Phylogenetic systematics, in ‟Annual review of entomology", 1965, X, pp. 97-116.
Hinton, H. E., Some structures of Insects as seen with the scanning electron microscope, in ‟Micron", 1970, I, pp. 84-108.
Jander, R., Insect orientation, in ‟Annual review of entomology", 1961, VIII, pp. 95-114.
Jeannel, R., La genèse des faunes terrestres, Paris 1942.
Jurato, S., Submicroscopic structure of the inner ear, Oxford 1967.
Kennedy, J. S., Behaviour as physiology, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edimburgh-London 1967, pp. 249-265.
Kennedy, J. S., The emergence of behaviour, in 14th International Congress of Entomology. Abstracts, Canberra 1972, p. 337.
Landa, V., Juvenile hormone analogues as chemosterilants, in 14th International Congress of Entomology. Abstracts, Canberra 1972, p. 232.
Locke, M., What every epidermal cell knows, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 69-82.
MacArthur, R. H., Wilson, E. Q., An equilibrium theory of insular zoogeography, in ‟Evolution", 1963, XVII, pp. 373-387.
MacKerras, I. M., Composition and distribution of the fauna, in The Insects of Australia, Melbourne 1970, pp. 187-203.
Maddrel, S. H. P., A diuretic hormone in Rhodnius prolixus Stoal, in ‟Nature", 1962, CXCIV, pp. 605-606.
Maramorosh, K., Arthropod transmissions of plant viruses, in ‟Annual review of entomology", 1963, VIII, pp. 369-414.
Mayr, E., Animal species and evolution, Cambride 1963.
Mills, R. R., Hormonal control of tanning in the American cockroach. II. Assay for the hormone and the effect of wound healing, in ‟Journal of Insect physiology", 1965, XI, pp. 1269-1275.
Mittelstaedt, H., Control systems of orientation in Insects, in ‟Annual review of entomology", 1962, VII, pp. 177-198.
Montalenti, G., Aldrovandi, in Dizionario biografico degli Italiani, vol. II, Roma 1960, pp. 118-124.
Moss, W. W., Hendrickson, J. A., Numerical taxonomy, in ‟Annual review of entomology", 1973, pp. 227-258.
Müller, J., Handbuch der Physiologie des Menschen, Koblenz 1840.
Novak, V. J. A., The juvenilehormone and the problem of animal morphogenesis, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 119-132.
Pardi, L., L'orientamento astronomico degli animali. Risultati e problemi attuali, in ‟Bollettino di Zoologia", 1957, XXIV, pp. 473-523.
Richards, A. G., The integument of Arthropods, St. Paul, Minn., 1951.
Richards, O. W., The theoretical and practical study of natural Insect populations, in ‟Annual review of entomology", 1961, VI, pp. 147-162.
Rudall, K. M., The chitin protein complexes of Insect cuticles, in ‟Advances in Insect physiology", 1963, I, pp. 257-313.
Rudall, K. M., Kenchington, W., Arthropod silk. The problem of fibrous proteins in animal tissues, in ‟Annual review of entomology", 1971, XVI, pp. 73-96.
Salked, E. H., Wilkes, A., The ultrastructure of Insect surface by scanning electron microscopy, in ‟The Canadian entomology", 1968, C, pp. 1-4.
Schmidt, W. J., Die Doppelbrechung von Karyoplasma, Zytoplasma und Metaplasma, Berlin 1937.
Shaw, J., Stobbart, R. H., Osmotic and ionic regulation in Insects, in ‟Advances in Insect physiology", 1963, I, pp. 315-399.
Silvestri, F., Compendio di entomologia applicata, 2 voll., Portici 1934-1951.
Slifer, E. H., The thin walled olfactory sense organs on Insect antennae, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 233-245.
Smith, D. S., The trophic role of glial cells in Insect ganglia, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 189-198.
Smith, K. S., Insect virology, New York-London 1967.
Smith, S. G., Cytogenetics of Insects, in ‟Annual review of entomology", 1960, V, pp. 69-84.
Sokal, R. H., Numerical taxonomy, in ‟Scientific American, 1967, CCXV, pp. 106-116.
Treherne, J. E., Axonal function and ionic regulation in Insect central nervous tissues, in Insects and physiology (a cura di J. W. L. Beament e J. E. Treherne), Edinburgh-London 1967, pp. 175-188.
Uvarov, B. P., Grasshoppers and Locusts, vol. I, Cambridge 1966.
White, M. J. D., Cytogenetics and systematic entomology, in ‟Annual review of entomology", 1957, II, pp. 71-90.
White, M. J. D., Cytogenetics, in The Insects of Australia, Melbourne 1970, pp. 72-82.
Wigglesworth, V. B., The principles of Insect physiology, London 1939.
Wigglesworth, V. B., The hormonal regulation of growth and reproduction in Insects, in ‟Advances in Insect physiology", 1964, II, pp. 247-336.
Wilkinson, C., Some uses of computers in taxonomic entomology, in 14th International Congress of Entomology. Abstracts, Canberra 1972, p. 91.
Wyatt, G. R., The biochemistry of sugars and polysaccharides in Insects, in ‟Advances in Insect physiology", 1967, IV, pp. 287-360.
Yokoyama, T., Sericulture, in ‟Annual review of entomology", 1963, VIII, pp. 287-306.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


